
Палеонтологические доказательства эволюции
Доказательства из области палеонтологии, как палеозоологии, так и палеоботаники, уже не имеют характера косвенного доказательства эволюционного процесса, а приносят нам непосредственные данные об эволюционном процессе. Дарвин приводя в "Происхождении видов" доказательства из палеонтологии, подчеркивал их фрагментаричный и неполный характер. Со времен Дарвина наши сведения по палеонтологии многократно возросли, однако мы должны отдавать себе отчет в том, что окаменелости живших в давние времена животных могли возникать лишь в совершенно исключительных условиях, и что огромные пространства материков еще почти совершенно не исследованы палеонтологами. Следует также помнить, что сохранившиеся остатки очень некомплектны, а это значительно затрудняет работу палеонтологов.
Прежде, чем мы кратко остановимся на вопросе, каким путем могли возникнуть окаменелости, следует хотя бы в общих чертах познакомиться с эпохами, которые историческая геология отмечает в истории нашей Земли. Большинство авторов считает, что жизнь на нашей планете возникла примерно два миллиарда лет тому назад. Геолог делит историю Земли на эры, эры на периоды, а периоды на эпохи.
На приведенной таблице схематически представлена история земной коры, в которой выделяем четыре основных этапа: предкембрийский период, палеозойскую эру или древнюю эру, мезозойскую или среднюю эру и кайнозойскую или новую эру. Ясно, что предпалеозойский период был более длительным, чем все последующие периоды, которые для наших целей можно рассматривать вместе, как предкембрийский период. На таблице проведена продолжительность отдельных эр, периодов и эпох.
Современная наука располагает методами, позволяющими довольно точно определить продолжительность отдельных эпох. Ученые воспользовались радиоактивностью некоторых элементов, как например, урана, который подвергается самопроизвольному распаду с постоянной скоростью, в результате чего образуется определенный изотоп олова. Изучая в горных породах отношение этого изотопа к урану, можно определить возраст горной породы, а тем самым и возраст найденных там окаменелостей.
Симпсон, стремясь наглядно показать взаимоотношение периодов времени, в которые различные формы жизни развивались на Земле, приводит следующий пример. Он, для большей образности, продолжительность всей жизни, от самого ее возникновения, принимает за сутки, то есть 24 часа. Это значит, что условные 24 часа соответствуют двум миллиардам лет в геологической шкале.
В этой шкале первые многочисленные окаменелости морских животных появляются примерно в 18 часов. В 20 часов начинают появляться первые сухопутные растения, а через полчаса встречаются окаменелости первых насекомых и земноводных. Период расцвета пресмыкающихся начался в 21 час 30 минут, а млекопитающих в 23 часа. Человек появился за одну минуту перед полуночью, а весь исторический период человечества продолжается не больше, чем 0.25 секунды.
ГЕОХРОНОЛОГИЧЕСКАЯ ИСТОРИЯ ЗЕМЛИ
Таблица 1
| Миллионы лет от начала | Продолжительность в млн. лет | Эры | Периоды | Эпохи | ||
| 0,25 | 75 | 1 | 0,25 | Кайнозойская | Четвертичный | Голоцен (Современная) |
| 1 | 1 | Плейстоцен | ||||
| 12 | 74 | 11 | Третичный | Плиоцен | ||
| 28 | 16 | Миоцен | ||||
| 39 | 11 | Олигоцен | ||||
| 58 | 19 | Эоцен | ||||
| 75 | 17 | Палеоцен | ||||
| 135 | 130 | 60 | Мезозойская | Меловой | ||
| 165 | 30 | Юрский | ||||
| 205 | 40 | Триасовый | ||||
| 230 | 300 | 25 | Палеозойская | Пермский | ||
| 280 | 50 | Каменноугольный | ||||
| 325 | 45 | Девонский | ||||
| 360 | 35 | Силурийский | ||||
| 425 | 65 | Ордовикский | ||||
| 505 | 80 | Кембрийский | ||||
| 2000 | 1500 | Предкембрийский период | ||||
Останки животных, вымерших в исторический или предисторический период, мы еще не относим к окаменелостям. Если же животные или растения жили больше, чем 100000 лет тому назад, то остатки их уже называются окаменелостями. Животные, которые погибли относительно недавно, могли в исключительных условиях хорошо законсервироваться. В этом случае мы находим не только твердые части скелетов, но также кожу, волосы и внутренние органы.
Так, например, во льдах Сибири сохранились замороженные тысячелетиями мамонты, а в Старуни, в бывшей Восточной Малопольше, в озокерите был найден прекрасно сохранившийся носорог, голова мамонта и другие животные, как птицы и насекомые. В этом случае экземпляры так хорошо сохранились, что на микроскопических срезах мышц носорога можно было заметить поперечную полосатость мышечных волокон. Этот носорог, набитый и смонтированный, является украшением Музея Института Зоологии Польской Академии Наук в Кракове (рис. 29).

Рис. 29. Шерстатый носорог из Старуни (плейстоцен). Коллекция Краеведческого Музея Польской Академии Наук в Кракове
В очень сухих гротах, в юго-восточных штатах Соединенных Штатов Америки и в Патагонии обнаружены хорошо сохранившиеся остатки огромных ленивцев, их кожа, волосы, сухожилия а также и их экскременты. Эти животные вели наземный образ жизни в отличие от современных, живущих на деревьях, и жили несколько тысяч лет тому назад (рис. 30).

Рис. 30. Реконструкция огромного ленивца из Южной Америки (плейстоцен); по J. August и Z. Burian.
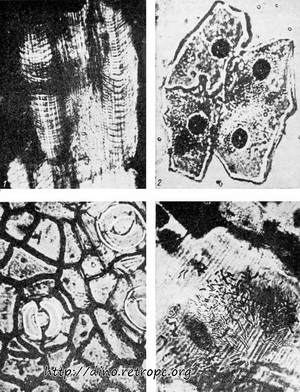
Рис. 31. Находки из Geiseital, сохранившиеся в слоях бурого угля. Прекрасное состояние этих находок позволило исследовать их микроскопическое строение. 1 - волокна поперечнополосатых мышц, 2 - эпителиальные клетки кожи лягушки, 3 - щели в кожице листа, 4 - бактерии в трахее хруща; по Z. Bulow
Однако такие находки очень редки относятся к сравнительно недавнему времени.
К гораздо более отдаленным временам относятся замечательно законсервированные в янтаре, то есть в смоле древних, деревьев, насекомые, паукообразные и части растений. Некоторые из этих окаменелостей насчитывают 50 и больше миллионов лет. Хорошей консервационной средой являлись болота и торф, находящийся в болотах, который насчитывает десятки миллионов лет. В таком виде в Австралии найдены хорошо сохранившиеся части деревьев, а в обнаруженных в Германии остатках животных можно было исследовать под микроскопом детали клеточного строения (рис. 31).
Целое животное может подвергнуться окаменению лишь в исключительно редких случаях. В таком виде найдены жабы и другие животные около Quercy во Франции, организм которых был насыщен солями фосфорной кислоты. Мягкотелые животные могли оставить отпечатки в мягкой породе, которые затем окаменевали. В таком виде сохранились, например, отпечатки медуз, тело которых, как известно, имеет желеобразную консистенцию. В других случаях от животных может остаться тонкий слой угля, который довольно точно отображает их первичную форму. Иногда же организм, затопленный в отложениях, подвергается разрушению, а оставшееся пустое пространство в дальнейшем может заполниться минеральными субстанциями, образуя, таким образом, его слепок.
Чаще всего, однако, то, что мы находим как окаменелости, это твердые части давних организмов. От позвоночных животных обычно остаются кости и зубы. Древняя твердая ткань, состоящая из солей угольной и фосфорной кислоты, остается на месте, а мягкие части разрушаются и после них остаются пустые пространства, которые затем могут заполниться минеральными субстанциями, чаще всего кремнеземом или углекислым кальцием. Поэтому ископаемые кости обычно весят гораздо больше свежевыпрепарированных костей, и могут быть окрашены в различные цвета.
Также и панцири животных, построенные из карбоната кальция, часто сохраняются неизменными. Иногда же материал панциря растворяется, а свободное пространство заполняется кремнеземом с образованием слепка. Хитиновые части насекомых, пауков и других построены из очень стойкого материала и могут сохраняться не изменяясь. У позвоночных животных иногда можно найти слепки черепа, заполненные минеральными субстанциями, тогда, как сами кости могут подвергнуться разрушению.
От давних организмов, живших в минувшие геологические эпохи, остались не только следы в виде частей их тела, главным образом костей и зубов. Сохранились отдельные стадии их развития, найдены, например, окаменевшие яйца пресмыкающихся - динозаров, окаменевшие экскременты, так называемые копролиты, и даже следы животных, оставшиеся на мягком грунте, который со временем преобразовался в сохранившиеся до настоящего времени горные породы.
На основании этих данных палеонтолог может составить себе представление не только о величии и форме вымерших видов животных, но и об их образе жизни, пище, которой они питались и так далее. По найденным костям животных можно реконструировать их мягкие части. Однако это трудная задача и реконструкция не во всех случаях заслуживает доверия. В настоящее время палеонтология обладает часто сложными методами извлечения окаменелостей из горных пород и соответствующей их консервации. Иногда палеонтолог приготавливает очень тонкие шлифы, чтобы ближе познакомиться со строением организмов и их частей.
С дидактической точки зрения все доказательства, которые нам представляет палеонтология, можно изложить в трех основных пунктах. На приведенном эскизе, позаимствованном у Симпеона, представлено увеличивающееся богатство разных форм животных и растений на протяжении очередных этапов истории нашего земного шара (рис. 32). По мере исследования окаменелостей от наиболее давних до все более близких нам времен, отмечаем увеличение количества типов животных и растений, увеличение количества форм и все более высокие ступени их организации.
Одновременно организмы приспосабливаются к жизни во все более новых средах. Вначале мы встречаем организмы, жившие исключительно в море, затем наступает заселение суши. Жизнь завоевывает все более новые среды и в этом выражается ее экспансия. Сухопутные растения и животные постепенно занимают области, отличающиеся разным климатом, разной влажностью, некоторые животные приспосабливаются к жизни в земле, другие приобретают способность к полету, третьи же приспосабливаются к жизни в пресных и соленых водах.
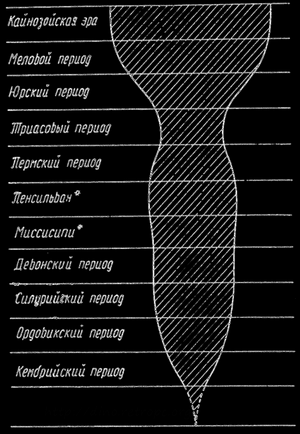
Рис. 32. Экспансия жизни; по Дж.Г. Симпсону. (* - соответствует каменноугольному периоду).
Приспособление к различным средам влечет за собой все большую специализацию организмов. Вид животных, который питается десятью разными видами растений, может эволюционно преобразоваться в десять новых видов, из которых каждый будет питаться только одним отдельным видом растений (Симпсон). Следует помнить, что экспансия жизни сама создает новые среды, которые в свою очередь могут быть заселены новыми видами. Так, например, вначале должны были появиться деревья, образующие леса, а затем эволюционным путем развились растения, называемые эпифитами, для которых деревья являются опорой. В свою очередь в зарослях этих эпифитов могут жить различные виды животных, приспособленные к этой новой среде.
На рисунке, изображающем экспансию жизни, видно, что в одном периоде, а именно, между пермским и триасовым, то есть на границе палеозойской и мезозойской эры, наступил как бы кризис. Уменьшилось разнообразие и богатство форм жизни. Это, однако, не длится долго (в геологическом понятии времени). Жизнь начинает с новой силой обогащаться и распространяться. Кроме того палеонтология учит нас о том, что жизнь в своих разнообразных формах никогда не достигает какого-то равновесия, постоянно наступают эволюционные изменения, постоянно развиваются новые формы, а многие формы гибнут. Ясно, что некоторые организмы могут иногда продержаться многие миллионы лет не изменяясь. Но это не противоречит общему принципу постоянных эволюционных изменений.
Наконец, изучая формы в различные геологические периоды, от наиболее старых, следует отметить, что эволюционные изменения ведут к прогрессу и более сложной организации организмов. Прогресс проявляется во все более совершенном приспособлении организмов к особым условиям их жизни, в изменениях их строения и функций, делающих возможным завоевание все новых сред, наконец, более высокая степень организации позволяет навязать лучший контакт с окружающей средой, отбирать большее количество стимулов и более совершенно реагировать на стимулы среды и ее изменения.
Если старые эволюционные теории ставили этот прогресс в зависимости от каких-то неопределенных свойств и сил, действующих в самих организмах, то теория естественного отбора объясняет естественным образом, почему в процессе эволюции организмы все более приспосабливались и почему уровень их организации поднимался на все более высокую ступень. Излагая теорию Ламарка, мы обращали внимание на то, что этот автор принимал принцип самоусовершенствования как имманентное свойство живой материи. Давнишняя мысль Ламарка оживала неоднократно в новой форме.
В недавние времена Бергсон, французский философ, приписывает силу эволюционного усовершенствования неизвестному фактору, а в последнее время палеонтолог, иезуит Teilhard de Chardin аналогичным образом старается объяснить задачу эволюционного прогресса. И лишь теория, впервые сформулированная Дарвином, объясняет этот прогресс, не прибегая к помощи факторов, недоступных науке.
Лучше всего эти общие выводы, выдвинутые палеонтологами, можно представить при помощи короткого наброска постепенного развития фауны в следующих за собой геологических эрах, периодах и эпохах. Появление все большего разнообразия форм, все более высокая организация их, является непосредственным и убедительным доказательством эволюционного принципа. Одновременно обратим внимание и на вторую группу доказательств, представленных палеонтологией. Эта наука открывает промежуточные формы, объединяющие отчетливо разделенные в настоящее время более крупные систематические категории. Современная палеонтология знает уже много таких промежуточных форм. Мы не можем заняться подробным их разбором и поэтому ограничимся лишь переходными формами между отдельными классами позвоночных. Позвоночных мы выбрали потому, что человек тоже является одним из них, и ознакомление с эволюционным развитием этой группы животных необходимо для познания всей нашей родословной. Наконец, описывая родословную позвоночных животных, мы сможем остановится также на третьей группе палеонтологических доказательств. Они относятся к так называемым палеонтологическим (филогенетическим) рядам. В отдельных случаях окаменелости какой-нибудь группы животных так обильны в следующих за собой геологических слоях, что на их основании можно как бы шаг за шагом проследить эволюционные изменения от очень древних предков до форм, живущих в настоящее время. Этим путем, например, удалось изучить родословные некоторых видов современных млекопитающих животных, например лошадей, от наиболее ранних предков. Так как нашей основной задачей является изучение эволюционного развития позвоночных, следует кратко представить принцип систематики этой группы животных (Collbert).
Позвоночные вместе с другими формами относятся к типу хордовых, Chordata. Характерным признаком всех хордовых является постоянное или временное наличие хорд, то есть спинной струны, и тубулярной нервной системы, то есть нервной трубки, расположенной дорзально по отношению к хорде. Кроме того передний отрезок пищеварительной системы исполняет дыхательную функцию (образование жаберных дуг).
Тип (phylum) хордовых (Chordata) зоологическая систематика делит на подтипы. Первым из них являются полухордовые (Hemichordata). Это морские животные червеобразной формы. В передней части тела их видны жаберные щели. Имеется дорзально расположенная нервная система и зачаток нотохорда.
Вторым подтипом хордовых являются личиночнохордовые (Tunicata) называемые также Urochordata. Это тоже морские животные. Личинки одной из групп этих животных имеют дорзальную нервную систему, спинную струну в хвостовой части, откуда и название Urochordata, и жаберные щели. Однако после того, как личинка оседает и преображается, многие из этих признаков подвергаются обратному развитию или исчезают.
Третий подтип хордовых носит название Cephalochordata, к которым относятся хорошо известный ланцетник (Branchiostoma), живущий в морях. Дорзальная нервная трубка у них хорошо развита. Однако спереди эта трубка не образует отчетливой отдельной мозговой части. Хорда остается на всю жизнь. У зрелых форм жаберные щели очень многочисленны и открываются в околожаберные пространства.
Наконец, последним подтипом хордовых являются позвоночные (Vertebrata), к которым относятся многочисленные формы животных, отличающиеся некоторыми общими признаками. Позвоночные имеют дорзально расположенную трубчатую нервную систему. Передняя часть мозговой трубки образует мозг. Нотохорд всегда имеется у зародышей но в дальнейшем чаще всего подвергается обратному развитию и вместо нее развиваются позвонки. Жаберные щели и лежащие между ними жаберные дуги или существуют всю жизнь и являются функциональными органами, или же появляются только у зародыша и не функционируют. Сердце расположено на вентральной стороне туловища. Глаза развиваются благодаря выпячиванию стенки мозга, которое образует зачатки зрительных луковиц. Подтип позвоночных зоологическая систематика делит на два надкласса (superclassis). Одним из них являются "рыбы", или водные позвоночные, другой образуют сухопутные позвоночные, то есть Tetrapoda (Colbert).
В свою очередь надкласс "рыб" (в широком значении этого слова) делится на четыре подкласса (Romer). Первый подкласс включает безчелюстные (Agnatha), из которых в настоящее время сохранились только миноги и миксины, живущие в воде. Личинки миног питаются микробами и органическими остатками, содержащимися в иле, после превращения во взрослую особь миноги ведут паразитический образ жизни, питаясь кровью рыб, к которым прицепляются. Хотя особый вид жизни миноги должен был вызвать в ней далеко идущие преобразования, в сравнении с ее примитивными предками, она все же сохранила очень много первичных признаков. "Когда мы смотрим на миногу, то как бы через мглу видим древних позвоночных, которые жили почти пол миллиарда лет тому назад" (Colbert).
Другой подкласс заключает давно вымершие панцирные рыбы (Placodermi). Это первые рыбы, которые имели челюсти. У бесчелюстных между жаберными щелями находим расположенные одна за другой жаберные дуги, построенные из элементов скелета. Первая из них называется челюстной дугой, другая - подъязычной. У панцирных рыб первая из этих дуг образует челюсти, отделенные от черепа и соединенные с ним только при помощи связок. Placodermi называются панцирными рыбами, так как они имели наружный костный панцырь, построенный из пластинок. Такое же оснащение имели многочисленные ископаемые безчелюстные рыбы, которых раньше также относили к панцирным рыбам. Placodermi известны только из окаменелостей.
Следующий подкласс рыб составляют хрящевые (Chondrichthyes). Они развились в морях, где живут до настоящего времени. Из живущих в настоящее время следует назвать акул и химер. Кроме интересных физиологических и биохимических особенностей (наличие в тканевых жидкостях большого количества мочевины), они отличаются полностью хрящевым скелетом.
Наконец, последним подклассом рыб являются костистые (Osteichthyes). Этот подкласс рыб включает очень многочисленные и разнообразные формы. Сюда относятся так называемые двоякодышащие рыбы. Эти рыбы и сейчас живут в Австралии, Южной Америке и Африке. Двоякодышащие рыбы могут дышать не только жабрами, но и "легкими", которые являются гомологом плавательного пузыря других рыб. При помощи жабр двоякодышащие рыбы дышат воздухом растворенным в воде, тогда как благодаря так называемым "легким", они могут дышать также атмосферным воздухом. У большинства же костистых рыб имеется плавательный пузырь, который служит рыбе для того, чтобы она могла удерживаться на соответствующей глубине в воде.
Наиболее многочисленной группой рыб среди Osteichthyes в настоящее время являются собственно костные рыбы, то есть Teleostei.
Наконец, очень большое значение с точки зрения эволюции имеют открытые недавно в море кистеперые рыбы, то есть Crossopterygii. Вплоть до 1939 г. считалось, что эти рыбы полностью вымерли около 75 миллионов лет тому назад. В 1939 г. выловлены живые представители этой группы - латимерии (Latimeria), вблизи восточного побережья Южной Африки. В настоящее время известно уже большее число представителей этого вида, выловленных вблизи Мадагаскара. Этот вид рыб заслуживает особого внимания потому, что именно он дал эволюционное начало следующему классу млекопитающих, а именно земноводным (Amphibia).

Рис. 33. Латимерия; по А.Ш. Ромеру
Среди этих последних различаем земноводных хвостатых, как тритоны и саламандры, и бесхвостых - жабы, лягушки и другие. Кроме того к земноводным относятся безногие и слепые земноводные, живущие в земле в тропической Азии, Африке и Америке, так называемые Apoda.
Следующим классом позвоночных являются пресмыкающиеся, Reptilia. Из 15 отрядов пресмыкающихся известных в ископаемом состоянии в настоящее время сохранилось только 4. Один из современных отрядов представлен лишь немногочисленными представителями единственного вида Sphenodom punctatum, живущего на небольших островах вблизи Новой Зеландии. Следующими отрядами являются: черепахи (Chelonia), крокодилы и аллигаторы (Crocodylia), ящерицы и змеи (Squamata).
Два последние класса позвоночных это птицы (Aves) и млекопитающие (Mammalia).
После этого краткого систематического обзора хордовых следует в общих чертах изложить развитие различных форм жизни от наиболее ранних геологических периодов. До недавнего времени палеонтологи стояли перед загадкой, разрешить которую было нелегко. От самого начала палеозойской эры, то есть от кембрийской эры, были известны многочисленные окаменелости различных видов морских животных, тогда как суша того времени была еще совершенно лишена жизни. В предкембрийском же периоде или совершенно не обнаружено никаких окаменелостей, или же они будили сомнения, имеем ли мы в этом случае действительно дело с остатками древних организмов. Высказывались различные предположения, которые должны были объяснить отсутствие отчетливых следов жизни в этих очень давних периодах, предшествовавших палеозойской эре, ни одно из них, к сожалению, не было достаточно убедительным.

Рис. 34. Предкембрийская фауна из Южной Австралии. 1 - Tribrachidium, форма, не напоминающая никакое из известных животных, 2 - Spriggina fluondersi, сегментированный червь, напоминающий живущих в настоящее время животных, 3 - Dickinsonia costata, возможно, что эта форма является родственной червям, 4 - Parvancorina minchami является представителем неизвестного до настоящего времени типа животных; по М.F. Glaesner
Лишь после II Мировой Войны в Южной Австралии были обнаружены богатые находки морской фауны, относящиеся к периоду примерно 600 миллионов лет тому назад, которые в какой-то степени были связаны с более давними открытиями в южной Африке и Англии. Животные, выброшенные волнами океана на берег, оставляли отпечатки в мягком иле, которые затем покрылись слоем песка. Этот песок в дальнейшем подвергся цементации раствором кремния, образуя, таким образом, точные слепки организмов, которые вначале находились в мягком иле.
До настоящего времени из австралийских раскопок собрано около 600 образцов. Среди них находили медуз, относящихся к вымершим родам, мягких кораллов, родственных живущим формам, таким, как Pennatula, сегментированных червей с головным щитом и другие формы двусторонне симметричных организмов, напоминающие некоторые червяки, и два загадочных организма, не напоминающие никаких из живущих в настоящее время животных. Найдены также формы, которые вероятно следует отнести к губкам и водорослям.
Возможно, что некоторые из найденных предкембрийских червей составляли группу, которые позже, в кембрийском периоде, дали начало примитивным вымершим членистоногим, называемым трилобитами. Несомненно, что в этом периоде имелись многочисленные одноклеточные животные и растения. В начале палеозойской эры, то есть в кембрийском периоде и следующем ордовикском, животные и растения жили исключительно в морях. Суша еще совершенно не была заселена.
Если сохранившиеся окаменелости раннего кембрия можно отнести только к восьми разновидностям животных, то в позднем ордовике имеется уже 13 разновидностей животных. Давние формы гибли, а новые занимали их места. В настоящее время сохранился лишь один род ордовикского периода - Ligula, относящийся к плеченогим (Brachiopoda). В морях ордовикского периода жили водоросли, возможно, что уже имелись грибы. Сосудистых растений еще не было. Из животных встречаем большое количество одноклеточных, губок, кишечнополостных, плеченогих, брюхоногих (Gastropoda), головоногих (Cephalopoda), характерной формой которых являлись организмы, родственные живущим в настоящее время корабликам (Nautilus). Одни из них были свернуты, напоминали Nautilus, другие были прямыми и достигали величины 5 метров. Зато современных головоногих, таких, как каракатицы и осьминоги, еще не было.
Из членистоногих жили вымершие позже трилобиты (рис. 35). Это были ракообразные, которые достигли вершины своего развития в ордовике. Из членистоногих следует также отметить представителей вымершей группы Eurypteridae, родственных скорпионам и паукам. В морях жили многочисленные формы типа иглокожих, к которым в настоящее время относятся морские ежи, морские звезды, голотурии, морские лилии и офиуры. В конце ордовикского периода появляются первые позвоночные, а именно, безчелюстные рыбы.
В связи с появлением первых позвоночных следует упомянуть о современных взглядах на эволюционное развитие предков хордовых и позвоночных животных. Некоторые авторы считают, что поскольку личинки иглокожих и полухордовых обладают характерными сходствами, следует принять, что между этими двумя группами животных имеется родственная связь. Полухордовые принято считать какой-то боковой ветвью в эволюционном развитии хордовых. Польский палеонтолог Козловски считает, что загадочная группа животных, так называемые Graptolithina, живших в ранних периодах палеозойской эры, связаны родственными связами с полухордовыми, и поэтому Козловски считает, что хордовые взяли свое начало именно от граптолитов.
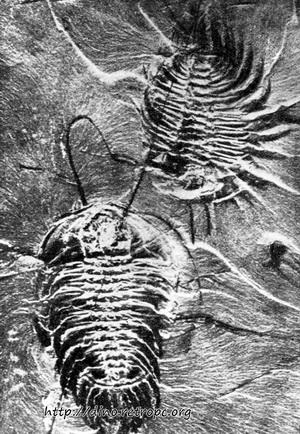
Рис. 35. Два представителя трилобитов с частично сохранившимися конечностями в кембрийском иле; по Р. Козловскому
Как видим, разные авторы вплоть до настоящего времени отстаивают различные точки зрения на происхождение типа хордовых. Мы также точно не знаем, какие формы примитивных хордовых дали начало позвоночным. Канадский ученый Berill считает, что позвоночные берут свое начало от аппендикулярий, относившихся к оболочникам (Urochordata). Из аппендикулярий эволюционным путем развилась свойственная им подвижная личинка, напоминающая головастика лягушки.
Berill считает, что эти подвижные личинки в мелких морских водах превратились в неотенические формы, то есть начали размножаться в личиночной стадии. Развивая эволюционным путем свои
В юрском периоде обнаружены четыре отряда первичных млекопитающих: Triconodonta, Symmetrodonta, Pantotheria и Multituberculata. Triconodonta, величина которых колебалась от величины мыши до величины кошки, вымерли в меловом периоде. Symmetrodonta вымирают уже в конце юрского периода, a Multituberculata первые растительноядные млекопитающие, достигают начала кайнозойской эры и тоже в конце концов вымирают, не оставляя потомства. Таким образом, из примитивных млекопитающих только Pantotheria вышли победителями и дали начало всем млекопитающим, вершина расцвета которых приходится на третичный период кайнозойской эры.
Уже в конце мелового периода Pantotheria дают начало двум группам млекопитающих, а именно, сумчатым и первым плацентарным млекопитающим (Placentatia). К меловому периоду относятся сумчатые - близкие родственники современных американских сумчатых крыс - опоссум, остатки которых обнаружены в Северной Америке. В Монголии также обнаружены ископаемые плацентарные млекопитающие, относящиеся к отряду насекомоядных и родственные современным землеройкам (Sozicidae).
Встает вопрос, почему так долго, от юрского периода до самого конца мезозойской эры, млекопитающие занимали столь незаметное место среди сухопутной фауны позвоночных. Чтобы объяснить этот факт, Симпсон приводит следующую гипотезу: "Когда появились земноводные, возник новый способ жизни, который не знал конкуренции. Наступила быстрая их экспансия, которую следовало ожидать. Так же и первые пресмыкающиеся имели широкий доступ к новым способам жизни, новые среды были почти лишены конкурентов. Благодаря этому они не только быстро развились, но и вытеснили земноводных из тех сред, которые те занимали до того времени. Когда же появились млекопитающие, положение было совершенно другим. Различные среды, которые могли быть заняты млекопитающими, и которые они в конце концов завоевали, были уже заняты пресмыкающимися".
"В общей экологической картине материков мезозойской эры млекопитающие являлись лишь малочисленными специализированными ветвями пресмыкающихся. Они занимали ограниченные экологические ниши и не имели выхода из них из-за натиска, который оказывали полные еще сил и успеха специализированные формы пресмыкающихся. Млекопитающие могли в этих условиях пережить юрский и меловой периоды вероятно лишь благодаря своему физиологическому совершенству и плодовитости. В этой борьбе за существование мозг немногим помогал первым млекопитающим. Из того, что нам известно, млекопитающие мезозойской эры были немногим интеллигентней пресмыкающихся".
В Скалистых Горах, рядом с остатками динозавров, найдены остатки мелких примитивных млекопитающих, относящиеся к позднему меловому периоду. Уже в самом начале третичного периода, а именно в палеоцене, динозавров уже совершенно нет, зато можно встретить представителей млекопитающих, относящихся к двум отрядам, а именно, сумчатых и насекомоядных. Вскоре, в том же геологическом периоде, мы встречаемся с бурным и быстрым развитием разных млекопитающих. Среди них сохранились окаменелости уже довольно крупных растительноядных копытных млекопитающих, плотоядных хищников, первых грызунов и других.
Совершенно ясно, что тогда произошло. Млекопитающие, лишенные конкуренции пресмыкающихся, которые массовым образом вымирают в конце мезозойской эры, занимают их места, приспосабливаясь к различным условиям и средам. У млекопитающих, теплокровных животных, наступило быстрое развитие мозга, благодаря чему они легче справлялись с различными жизненными трудностями. Отличаясь живородностью, они, после того, как большинство пресмыкающихся погибло, могли не только занять их места, но и завоевать те среды, которые были недоступны пресмыкающимся.
Так же, как в прошлом пресмыкающиеся из немногочисленных архаических форм дифференцировались в различные ветви, приспособленные к жизни в разных средах, так теперь млекопитающие, выводящие свой род из невзрачных предков, завоевывают в кайнозойской эре все новые среды и приобретают различные приспособления. Если какая-то группа дифференцируется в многочисленные подгруппы, отличающиеся строением, функцией и способом жизни, то мы говорим об адаптивной радиации. Опираясь на исследованиях палеонтологов, следует принять, что экспансия млекопитающих и их адаптативная радиация наступила не в результате вытеснения пресмыкающихся примитивными млекопитающими, а в результате того, что уже раньше; по неизвестным до настоящего времени причинам, наступило быстрое вымирание фауны пресмыкающихся.
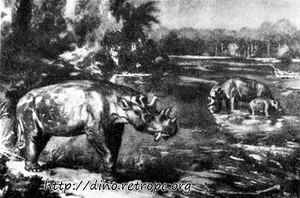
Рис. 49. Uintatherium, остатки этого млекопитающего найдены в слоях эоценовой эпохи в Северной Америке; по J. August и Burian
В третичном периоде наступила быстрая модернизация примитивных млекопитающих. Вероятно уже в олигоценовой эпохе все современные отряды млекопитающих имели своих представителей. Однако как в эоцене, так и в олигоцене млекопитающие выглядели иначе, чем современные животные. В то время жили еще представители многих вымерших в настоящее время групп, а кроме того многие предки современных видов иначе выглядели, чем их более поздние потомки. Некоторые из таких древних млекопитающих представлены на приведенных рисунках.
Следует также помнить, что процесс модернизации млекопитающих животных не охватил всех континентов. Австралия, отделенная от других континентов, имела, по существу лишь представителей сумчатых, которые претерпели адаптивную радиацию в различных направлениях. Так же и на континенте Южной Америки, отделенном от Северной Америки от начала до самого конца третичного периода, существовали соответствующие условия для адаптивной радиации млекопитающих животных.
В кайнозойской эре можно часто наблюдать следующее явление: животные, вначале небольших размеров, постепенно увеличиваются и достигают иногда огромных размеров. Со сходным явлением мы также встречаемся при эволюции пресмыкающихся в мезозойской эре.
Останавливаясь на эволюции млекопитающих в кайнозойской эре, мы можем указать на третью группу эволюционных доказательств, представленных палеонтологией. В некоторых случаях количество найденных окамёнелостей в очередных геологических формациях так велико, что на их основании можно шаг за шагом проследить постепенное преобразование определенных форм, от очень отдаленных во времени предков до современных видов. Лучше всего разработан эволюционный ряд лошади.
Вся истинная эволюция семейства лошадиных происходила в третичном периоде на территории Северной Америки. В наиболее раннем третичном периоде, в палеоцене, первично растительноядные копытные животные (Condylarthra) и примитивные хищники (Creodonta) находились в родственных отношениях. К Condylarthra относился Phenacodus, передние и задние конечности которых имели по пять пальцев с небольшими копытами. Судя по форме зубов, эти животные питались как мясной, так и растительной пищей. Ввиду того, что до настоящего времени не обнаружено промежуточных звеньев между этой формой и первыми формами, отнесенными к копытным, Phenacodus в систематике включен в семейство Condylarthra, представители которого вымерли в течение следующей эпохи, эоцена.
В эоцене имелся уже первый представитель отряда копытных, семейства Equidae который называется Eohippus. Это животное однако скорее напоминало Phenacodus, чем современную лошадь. Это было небольшое животное, величиной с довольно крупную собаку или лисицу. На передних конечностях оно имело по четыре, а на задних по три пальца, законченных копытами. Зубы его свидетельствуют о том, что это животное еще не могло питаться травой, а пища его состояла из сочных листьев, почек, плодов и семян. Оно жило, как и Phenacodus в Северной Америке, которая в то время отличалась очень теплым и влажным климатом. Это небольшое животное жило в лесу и питалось буйной сочной растительностью.
В настоящее время известно большое количество ископаемых форм, которые связывают этого первого представителя Equidae с современной лошадью. Однако ошибочным было бы считать, что вся эволюция, от этих наиболее давних до современных форм, протекала в одном направлении, вдоль одной прямой линии предков, путем постепенного увеличения размеров тела, постепенного уменьшения количества пальцев и изменения зубов, в результате чего животное приспособилось к питанию сухими травами степей и саванн.
В родословной лошадиных отличаем не одну, а большое количество линий развития. Пытаясь представить эволюционное развитие лошади, мы были бы ближе всего к правде, если бы представили себе этот процесс в виде разветвленного дерева, многочисленные ответвления ствола и ветви которого заканчиваются слепо, не давая новых ветвей и представляя вымершие линии, которые не оставили потомства. Замечательный знаток эволюции этой группы животных, Симпсон, пишет следующее: "Увеличение размеров тела и изменения формы ног не происходили непрерывно, а спорадически. Величина всего тела изменилась мало в течение первых 15 и последних 5 миллионов лет. Увеличение тела не происходило равномерно и постоянно.

Рис. 50. Eohippus; по J. August и Z. Burian
Начиная от эоцена, предки лошадей дали начало трем главным и различным линиям развития, если принять во внимание строение и функцию конечности. Из этих трех первичных линий до настоящего времени сохранилась только одна. Сходным образом происходило эволюционное развитие всех других форм, которые мы можем проследить в палеонтологических рядах благодаря тому, что сохранилось большое количество ископаемых форм из разных геологических периодов.
В течение эоцена Eohippus дал начало многим видам и родам, которые жили как на своей родине, то есть в Северной Америке, так и перекочевали в Евроазию, так как эти материки в то время были соединены между собой. Однако те виды, которые поселились в Евразии, вымерли. В Америке в олигоцене распространился новый вид, Mesohippus, представители которого были крупнее, чем Eohippus, и имели на передних конечностях только три пальца. Одновременно, на что указывают результаты изучения черепов, наступили изменения и в мозге, полушария мозга значительно увеличились.

Рис. 51. Mesohippus; по J. August и Z. Burian

Рис. 52. Merychippus; no August и Z. Burian
В миоцене Mesohippus дал начало многочисленным видам, из которых два, а именно, Anchitherium и Hypohippus, снова заселили пространства Старого Света. Оставшийся в Америке Meryhippus подвергается важным эволюционным изменениям. В первую очередь изменяются зубы, поверхность которых все больше напоминает поверхность зубов современных лошадей. Эта эволюционная перемена была связана с изменениями климата Северной Америки в миоценской эпохе. Климат становился все более холодным и сухим, в результате чего все большие пространства занимали травянистые степи. Всякие наследственные изменения, которые помогали использовать растительную пищу прерий и саванн, закреплялись в процессе естественного отбора.
Поэтому не случайно именно в миоцене появились предки современной лошади, которые были все лучше приспособлены к питанию травянистой пищей. Поэтому нет ничего удивительного в том, что эволюционные потомки Meryhippus находят хорошие условия для своего дальнейшего развития, и в следующей эпохе, плиоцене, мы находим почти исключительно формы, происшедшие от предков типа Meryhippus. Одной из них был Hipperion, который из Северной Америки проник в Европу, Азию и даже Африку. Величиной с пони, Hipperion имел еще по три пальца на передних и задних конечностях.
В то же время в Америке развивается следующий вид, Pliohippus, который имел уже только один палец. Pliohippus вероятно является предком рода Equus, к которому кроме лошадей относятся еще зебры и ослы. В ледниковую эпоху, плейстоцене, род Equus, который возник в Америке, распространился в Евразию, Африку и Южную Америку, которая в то время уже была соединена с Северной Америкой Панамским перешейком.
Когда в Америке поселились ее первые жители, индейцы, которые прибыли из Азии, они встретили здесь лошадей, в большом количестве заселявших американские прерии. Когда же Америка была открыта белым человеком, на этом континенте уже не было лошадей. Лишь позднее туда были завезены лошади. До настоящего времени совершенно неизвестно, что явилось причиной гибели лошадей на их первоначальной родине, в Америке, ни одна из гипотез по этому вопросу не является достаточно убедительной. Неизвестно также, почему главная эволюция лошади происходила в Америке, почему те из животных, которые попали в Евразию, вымирали, не давая эволюционного потомства.
По мнению зоогеографа Дарлингтона, род Equus представлен в настоящее время семью видами, живущими в диком состоянии в Азии и Африке. Дикая лошадь, называемая лошадью Пржевальского, живут еще в некоторых частях Монголии и Китайском Туркестане. Другая дикая лошадь, тарпан, полностью исчезла, последний представитель этого вида был убит около 1860 года, в степи, к северу от Черного моря.
Кроме того, от Монголии и Тибета до Сирии встречается так называемый азиатский осел, или онагр, тогда как истинные дикие ослы (Equus asinus) встречаются в некоторых районах Африки. Кроме того в Африке живет еще четыре вида зебр.
Осел был приручен 4000 лет до нашей эры или еще раньше. В то же время, или несколько позже, была приручена лошадь. Первые исторические источники говорят о приручении лошади в двух отделенных частях Азии, а именно, в Месопотамии и Китае. Некоторые авторы предполагают, что в Месопотамии был приручен тарпан, тогда как китайские лошади скорей всего происходят от лошади Пржевальского.
Возможно, что лошади приручались неоднократно, как в Азии, так и Европе (Добзхански). Римляне, во время своих военных походов, встречали многочисленных диких лесных лошадей в Западной Европе. Однако неизвестно, насколько эти лошади отличались от тарпана или лошади Пржевальского. Вероятно все дикие лошади не относились к отдельным видам, а составляли лишь различные географические расы, которые могли скрещиваться, давая плодовитое потомство.
Со времени приручения лошади, человек путем естественного отбора, используя комбинативную изменчивость и появляющиеся от времени до времени мутации, мог вывести разнообразные расы, приспособленные для различных целей. Маленькие пони, крупные упряжные лошади, беговые лошади, являются лишь особыми расами и не заслуживают названия отдельных видов.
Как мы уже указывали выше, современная палеонтология знает много примеров палеонтологических рядов, сходных с рядом семейства лошадиных, которые дают нам возможность проследить постепенные эволюционные изменения, приведшие к формам современным или вымершим в недалеком прошлом. Хорошим примером может служить палеонтологический ряд слонов (Proboscidea).
Первичной формой принято считать род Moeritherium, живший в Египте в позднем эоцене. Это были животные величиной со свинью, с толстыми массивными ногами и плоскими копытами. В течение развития разных линий этого семейства отмечаем увеличение размеров тела, так что в конце концов все представители слонов относятся к истинным великанам. Одновременно несоразмерно увеличивается череп, шея укорачивается и удлиняется нижняя челюсть. Верхняя челюсть и нос вытягиваются, а со временем нос образует подвижной хобот. Вторая пара резцов разрастается и образует так называемые бивни.
Эволюционное развитие слонов идет в двух основных направлениях. Представители одного направления, Deinotherioidea, полностью вымирают в плейстоцене. К ним относился род Deinotherium, живший в Евразии и Африке. Под конец своего существования это были самые крупные животные из группы слонов. В нижней челюсти они имели два бивня, загнутые вниз, вероятно имели хорошо развитый хобот (рис. 53). Ко второму направлению относятся формы из группы Elephantoidea.

Рис. 53. Deinotherium giganteum, хоботный из Румынии (плейстоцен); по J. August и Z. Burian
Вскоре после того, как Moeritherium вымерли, в Египте появились более высоко развитые формы. Уже в раннем олигоцене первые мастодонты были гораздо крупнее своих предков Moeritherium, с относительно длинными ногами, причем строение их черепа указывает на хорошо развитый хобот. Два массивных дивня нижней челюсти торчали горизонтально. Один из родов Palaemastodon величиной достигал слона средних размеров.
Следующие окаменелости относятся к позднему миоцену. Это были уже очень крупные животные, некоторые из них во время плейстоцена проникали из Северной в Южную Америку. У одной из групп бивни были изогнуты вниз, у других представителей, обитавших в Азии и Северной Америке, бивни были очень широкие, в виде лопаты, и вероятно служили для выкапывания растений со дна мелких вод.
Исследователи предполагают, что из первичного семейства Palaeomastodon развились также формы с короткой челюстью, почти совершенно без бивней в нижней челюсти. К ним относится Mastodon americanus с сильно изогнутыми верхними бивнями. Он не достигал величины современных слонов. Он жил в Северной Америке до конца плейстоцена, а сохранившиеся мелкие части его указывают на то, что он был покрыт длинными ржавобурыми волосами. Исследования, произведенные при помощи радиоактивного углерода С14, показали, что эти мастодонты жили еще около восьми тысяч лет тому назад. Они вымерли после того, как Америку заселили азиатские народы, за несколько тысяч лет перед тем, как Америка была открыта белым человеком.

Рис. 54. Огромный вымерший торфяной олень из Ирландии (плейстоцен); по A.S. Romer
Следующая линия происходит от тех форм слонов, которые жили в миоцене. К ним относится род Stegophodon. Нижняя челюсть у этих животных была укорочена, а верхние дивни были очень сильно развиты.
Следующим представителем является Stegodon - первый настоящий слон. Эти животные жили в Старом Свете начиная от позднего плиоцена, а главным образом в плейстоцене. Это были крупные животные с длинными изогнутыми верхними бивнями, тогда как нижняя челюсть была укорочена и лишена бивней. Из группы Stegodon в последствии развились мамонты и современные слоны.
Мамонты жили в эпоху плейстоцена в Евразии, Африке и Северной Америке, куда попали из Старого Света. Это были огромные животные, разделенные на многочисленные виды. Одни из них были родственниками современных азиатских слонов (Elephas), другие же - современных слонов, живущих в Африке (Loxodontia).
Как пишет Colbert: "Человек жил одновременно с мамонтами и слонами в течение своего эволюционного развития. Для примитивного человека, живущего в Евразии и Африке, гигантские мамонты должны были быть страшными чудовищами, перед которыми было трудно спастись. Однако с прогрессом эволюции человека, когда он стал создателем орудий и оружия, он стал и ловцом мамонтов. Многочисленные находки в Европе указывают на то, что человек каменного века охотился на мамонтов, побеждая их хитростью и хорошо обдуманной стратегией. Мамонтов часто загоняли в глубокие рвы, где их убивали камнями, или заманивали в смертоносные ловушки. А когда человек проник из Азии в Новый Свет и нашел там мамонтов, то приступил к охоте на них и их уничтожению. Имеются многочисленные доказательства того, что первые жители Америки охотились на мамонтов вероятно восемь - десять тысяч лет тому назад".
Палеонтологические доказательства, как мы уже указывали выше, являются прямыми доказательствами эволюционного прогресса. Они свидетельствуют не только о постепенном развитии жизни от примитивных форм к формам, все лучше воспринимающим раздражения из окружающей среды, все выше организованных, но знакомят нас с переходными формами между отдельными группами животных и растений, в настоящее время отчетливо отделенных друг от друга; наконец, в палеонтологических рядах, показывают постепенную эволюцию от очень давно живших предков к современным формам.
Наука палеонтология, истинное развитие которой датируется лишь от выступления Дарвина, в большой степени подтвердила и дополнила те данные, которые старые эволюционисты добыли на основании анатомических и эмбриологических исследований. Сравнительный анатом или эмбриолог мог привести лишь косвенные доказательства, свидетельствующие о биологической эволюции. По сходству строения и развития он делал выводы о родстве между разными группами организмов и об их общем происхождении. Палеонтологические данные не только подтвердили чаще всего правильные взгляды анатомов и эмбриологов, но, кроме того, обнаружили многие формы, связывающие различные группы животных и растений; иногда они дают нам картину очередных, наступивших друг за другом, эволюционных перемен.