
Биогеографические доказательства эволюции
Доказательства из биогеографии относятся к распределению живых организмов в пространстве, то есть географического размещения животных и растений. Ввиду этого в биогеографии мы различаем зоогеографию и фитогеографию. Мы ограничимся лишь зоогеографическими данными. Зоогеографические данные еще и потому представляют особый интерес, что именно они главным образом, навели Дарвина на мысль о биологической эволюции. Данные из области биогеографии тесно связаны с палеонтологическими данными.
Дарвин, исследуя современную ему фауну Южной Америки, заметил, что там живут примитивные млекопитающие - ленивец и броненосец, которые обнаружены и в ископаемом виде, хотя эти последние и выглядят иначе, чем виды, живущие в настоящее время. Таким образом, уже со времен Дарвина начали сочетать зоогеографические и палеонтологические искания, и общим усилием биогеографов и палеонтологов получено множество убедительных доказательств эволюции живой природы. Однако из дидактических соображений мы займемся в данной главе исключительно фактами, почерпнутыми из области биогеографии, останавливаясь на разборе данных из палеонтологии в одной из следующих глав.
При изучении данных относительно размещения животных и растений, мы приходим к убеждению, что лишь на основании принципов эволюции и изменений, которые претерпели материки и моря в истории земной коры, можно эти данные объяснить логическим образом. Дарвин и его единомышленники, провозглашая эволюционизм и выступая против креационизма, подчеркивали, что если отбросить принципы эволюции, то размещение живых существ, которое имеет место в настоящее время, станет совершенно нам понятным, и будет представлять собой лишь комплекс хаотических, не связанных друг с другом фактов.
Можно было бы предполагать, что условия, господствующие на данной территории, решительным образом влияют на растительный и животный мир. Несомненно, что животные, живущие в тропических лесах Африки, Америки и Австралии обладают некоторыми общими чертами, как и животные, живущие в высоких горах, Арктике или пустыне. Глубинные морские животные так же обладают общими специальными свойствами, как, например органами вырабатывающими свет. Среди животных, живущих в пещерах, часто встречаем формы, лишенные глаз или с рудиментарными глазами. Эти животные могут относится к разным систематическим группам (насекомые, рыбы, земноводные), что свидетельствует о том, что особые световые условия пещер облегчает выживание там форм с рудиментарными глазами, появляющихся как внезапные наследственные изменения, то есть мутации. Эти же мутации в других условиях не имеют шансов победить в конкуренции с формами, обладающими нормальными глазами, и гибнут, тогда, как в темных пещерах могут с успехом жить.
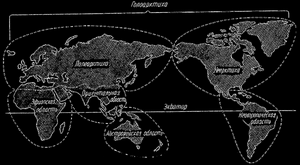
Рис. 27. Биогеографические области; по ДЖ. Симпсону и др.
Несмотря на то, что животные, живущие в одинаковых географических районах и в одинаковых условиях могут обладать определенными общими приспособлениями, нельзя считать, что эта конвергенция признаков свидетельствует об их общем происхождении. Поэтому, хотя у животных тропических лесов Африки, Америки и Австрии или африканских и австралийских бушах нередко можно обнаружить конвергенцию признаков, свидетельствующую о сходных условиях жизни, эти животные являются разными и зоологу не представило бы никакого труда различить их. Также обстоит дело с фауной и флорой на островах океана. Фауна островов Галапагосского архипелага, лежащего на запад от берегов Южной Америки, совершенно другая, чем фауна островов Зеленого Мыса, расположенных на запад от Африки, хотя климат и характер почвы в обоих случаях очень сходны.
Таких примеров можно было бы привести гораздо больше. Поэтому зоогеографы и фитогеографы в установлении биогеографических районов не руководствуются сходством климата или условий жизни вообще, а другими данными. Они принимают во внимание разницу в фауне определенных районов.
На основании размещения млекопитающих, а также птиц и других позвоночных, и, наконец, характерных признаков флоры, можно выделить следующие биогеографические области: Голарктическую, Эфиопскую, Восточную, Неотропическую, Мадагаскарскую, Антарктическую и Австралийскую. Некоторые авторы делят все материки только на три основные области, а именно: Арктогея, Неогея и Нотогея, причем Нотогея охватывает Австралию с соседними островами, Неогея - Южную и Латинскую Америку, тогда как все другие материки входят в состав Арктогеи.
Это последнее деление указывает на очень своеобразный характер фауны и флоры как Австралии, так и Южной Америки в сравнении с фауной и флорой других континентов. Это своеобразие Неогеи можно, как мы увидим, объяснить, принимая во внимание как эволюционные изменения, так и изменения, происшедшие при формировании континентов в процессе преобразования земной коры.
Придерживаясь первого из двух приведенных способов деления континентов на зоогеографические области, мы коротко приведем их характеристику. К голарктической области относится Северная Америка, арктические районы, Европа и северная часть Африки, а также Азия за исключением ее южной части. Эту область мы делим на Неарктику, то есть Северную Америку, и Палеарктику, то есть Европу, северную часть Африки и Азию без ее южной части.
В арктических областях живут следующие млекопитающие: мускусный бык, белый медведь, моржи. В Северной Америке встречаются лоси, северные олени, медведи, опоссумы, маралы, бизоны, в Европе - медведи, зубры, лоси, косули, в Азии - яки, лошади Пржевальского, двугорбые верблюды, тигры. В северной части Африки живут также крокодилы, некоторые узконосые обезьяны и страусы, которые встречаются также в Аравии.
К Эфиопской области относится Африка к югу от Сахары. Среди представителей фауны этой области можно назвать следующих: африканские слоны, жирафы, бегемоты, носороги, львы, зебры, узконосые обезьяны, гориллы, шимпанзе, страусы, полуобезьяны. В Восточной области, куда относится Индия, юго-западная часть Азии и Малайя, встречаются: индийские слоны, тапиры, полуобезьяны, орангутанги, азиатские носороги, тигры и узконосые обезьяны. В лесах Мадагаскара имеется большое количество лемуров. В Неотропической области, включающей Южную Америку, среднюю и южную часть Мексики, живут ягуары, американские тапиры, широконосые обезьяны, ламы, броненосцы, муравьеды, ленивцы, а из птиц-туканы, колибри, страусы нанду.
Наконец к Австралийской области относятся Австралия, Новая Гвинея, Новая Зеландия, Тасмания, Полинезийские и Гавайские острова. Из млекопитающих здесь живут очень примитивные клоачные млекопитающие, а именно: австралийский утконос и ехидна, живущие в Австралии и на Новой Гвинее. Кроме того в Австралии имеется большое количество представителей сумчатых животных, как например, кенгуру, а из птиц - страус эму и казуары. На Новой Гвинее живут казуары и райские птицы, на Новой Зеландии - бескрылые птицы киви, а из пресмыкающихся - очень примитивная форма - гаттерия (Sphaenodon). В Антарктике из млекопитающих встречаются тюлени, а из птиц характерны пингвины.
Приведенный выше краткий перечень главнейших животных, живущих в разных географических областях, облегчает нам более подробный разбор тех вопросов, которые имели особенно большое значение для развития эволюционной мысли. Как мы уже указывали выше, особенно своеобразную фауну и флору имеет Австралийская и Неотропическая области, хотя эта последняя соединяется с континентом Северной Америки.
Рассматривая фауну Южной Америки мы видим в ней две главные группы. К одной из них относятся примитивные млекопитающие, которые нигде больше не встречаются. Это муравьеды, ленивцы и броненосцы. Другая группа фауны родственна формам, живущим в Северной Америке. К ней относятся хопяки, олени и другие. Хотя современная Америка составляет один континент, так как северная и южная части ее соединены, фауна и флора Южной Америки так отлична от Северной, что была выделена в отдельную зоогеографическую область. Зато фауна Северной Америки имеет очень много общих черт с фауной Северной и Средней Азии и Европы. Поэтому обе области, то есть Неарктическую (Северная Америка) и Палеарктическую (Азия и Европа, то есть Евразия) мы объединяем вместе как Голарктику.
Приведем несколько примеров, иллюстрирующих сходство Неарктической и Палеарктической фауны. Лось, живущий в Европе, является очень близким родственником американского лося, олень из Северной Америки (Cervus canadensis) очень похож на европейского оленя (Cervus elaphus). Европейские северные олени являются близкими родственниками американских оленей карибу. Наш европейский зубр так близок американскому бизону, что в результате скрещивания этих двух видов можно получить плодовитое потомство.
Каким образом, на основании эволюции и изменений континентов можно себе представить с одной стороны сходство фауны Евразии и Северной Америки, а с другой - различие фауны обеих Америк?
Если в каком-то районе возникнет новый вид или группа животных, то по принципу эволюции она может постепенно распространяться от центра своего возникновения. На пути постепенного распространения форм, обладающих большими способностями приспособления к условиям окружающей среды и выходящих победителями в борьбе за существование, имеются, однако, непреодолимые преграды, в виде барьеров, тормозящих эмиграцию в новые районы. Для млекопитающих животных, которые мы, в основном, принимали во внимание, наиболее эффективными барьерами являются водные.
В настоящее время Северная Америка отделена морем (Беринговым проливом) от континента Евроазии. Однако когда-то было иначе. В течение всего третичного периода, то есть в течение 60-70 последних миллионов лет, когда особенно интенсивно путем эволюции развивались млекопитающие животные, Северная Америка не раз была соединена сушей с Евразией, а может быть и непосредственно с Европой. Благодаря этому могла происходить миграция животных из Евразии в Северную Америку и обратно. Особенно интенсивная миграция происходила из Евразии в Северную Америку.
Легко себе представить каким образом такое взаимное перемещение видов животных вызывало далеко идущие изменения в родимой фауне. Прибывающие новые формы приводили в расстройство общий баланс природы, создавали новые конкурентные связи. Наступающие изменения приводили к вымиранию целого ряда видов, а изменение направления в котором происходил естественный отбор вызывало новые эволюционные изменения.
Однако не все животные, населявшие Евразию, сумели проникнуть сухопутным путем в Северную Америку. По мере охлаждения климата в конце третичного периода на континент Северной Америки могли проникать лишь животные, приспособленные к жизни в более холодном климате. В еще большей степени на миграцию разных форм повлияла ледниковая эпоха, во время которой огромные территории северного полушария были покрыты массой льда, надвигавшегося с севера.
Таким образом, далеко идущее сходство фауны Неарктики и Палеарктики можно объяснить сухопутным соединением, существовавшим между Северной Америкой и Евразией, а также эволюцией форм, населяющих эти континенты, и прибывших в процессе миграции. Ввиду того, что Северная Америка относительно недавно отделилась от Евразии и теперь их разделяет морской барьер, то времени на значительные эволюционные изменения американской фауны с одной стороны и европейской и азиатской с другой, было недостаточно.
Иначе выглядит дело, если обратимся к Северной и Южной Америке. Обе части Америки до начала третичного периода были соединены между собой. В этом периоде, однако, единственными млекопитающими животными были примитивные формы, заселяющие как Северную, так и Южную Америку. Однако уже вначале третичного периода Северная Америка отделилась от Южной. Условия эволюционного развития Северной и Южной Америки становятся совершенно различными.
В Северной Америке наступает обмен между неарктической и евразийской фауной, тогда как в Южную Америку большинство новых форм, которые появляются на территории Северной Америки и прибывают сюда из Евроазии, не могут попасть через разделяющее их море. В результате этого в Северной Америке остаются только старые формы архаических млекопитающих, которые, развиваясь эволюционным путем, дают современных представителей примитивных животных этого района, как, например, броненосцы, ленивцы, муравьеды и так далее. (Это представление неверно. Большинство архаичных видов проникло в Центральную и южную часть Северной Америки после соединения Северной Америки с Южной в конце плиоцена. Таким образом, современные североамериканские формы броненосцев, муравьедов, ленивцев, опоссумов представляют собой недавних выходцев из Южной Америки (прим. редактора).)
Так как между Северной и Южной Америкой в то время были, однако, рассеяны небольшие острова, некоторые животные могли случайно проникнуть и Южную Америку. Таким образом, например, мы объясняем себе переселение представителей отряда приматов, из которых в Южной Америке развились широконосые обезьяны Нового Света.
Однако в конце третичного периода обе Америки снова соединяются Панамским перешейком. Несмотря на то, что этот перешеек, соединяющий обе Америки, неширок и покрыт густыми тропическими лесами, наступает миграция некоторых животных с севера на юг и с юга на север. Более высоко развитые животные родом из Северной Америки как например: тапиры, олени, являются тем вторым прогрессивным элементом в фауне Южной Америки, который способствовал вытеснению многих первичных копытных животных, свойственных Южной Америке. Лама в настоящее время является животным, типичным для Неогеи, но лишь с конца третичного периода, перед этим она жила в Неарктике. Это животное мигрировало в Южную Америку после того, как она соединилась с Северной, и целиком вымерло на своей первоначальной родине.
С другой стороны, некоторые примитивные млекопитающие животные, первоначальной родиной которых была Южная Америка, мигрируют на север. К таким относятся броненосцы, которые являются приобретенным архаическим элементом для Неарктики. Следует отметить, что эти выводы базируются не только на исследованиях фауны обеих Америк и на геолого-географических данных, но и на данных палеонтологии. Факты, которые дают эти дисциплины, соответствуют друг другу и взаимно дополняются, если одновременно принять принцип биологической эволюции.
Из всех зоогеографических и фитогеографических областей особенно своеобразной фауной и флорой отличается австралийская область, иначе Нотогея. Суша этой области изолирована от других материков. Мнение о том, что в раннем третичном периоде существовала связь между Южной Америкой и Австралией, кажется нам ошибочным. Только некоторые авторы предполагают, что Антарктида служила мостом между Южной Америкой и Австралией. Однако мы не можем полностью исключить такой возможности в более ранних геологических эрах, например, в мезозойской эре.
В настоящее время континент Антарктиды покрыт толстым слоем льда, однако открытые залежи угля указывают на то, что в мезозойской эре климат этого района был умеренным и там могли развиваться сухопутные животные. Но так как большинство исследователей считает, что уже от начала третичного периода Австралия была отделена от Антарктиды, то следует предположить что живущие в Австралии млекопитающие животные и птицы, а также и многие другие организмы должны были туда попасть через острова, лежащие между юго-восточной Азией и Австралией.
Из млекопитающих животных здесь живут, как мы уже указывали, представители клоачных, то есть яйцеродные млекопитающие, обладающие сходными признаками с пресмыкающимися и сумчатыми. Последние являются доминирующей группой автохтонных животных Нотогеи, приспособленных к различным условиям жизни. Клоачные вероятно представляют отдельную линию пресмыкающихся, развившуюся в направлении млекопитающих животных относительно недавно, не раньше, чем в конце третичного периода.
Сумчатые, же являются примитивными млекопитающими, которые появились уже в конце мезозойской эры.
Первичные сумчатые попали через острова на австралийский континент, уже совершенно отделенный от других материков. Не встретив тут большого количества других конкурентов, они развились эволюционным путем в различных направлениях, имитируя представителей различных отрядов млекопитающих, живущих на других континентах. Такое развитие в разных направлениях, позволяющее использовать различные условия существования, мы называем адаптивной радиацией. Таким образом, в Австралии можно встретить насекомоядных сумчатых, похожих на крота, сумчатых, похожих на муравьеда, белку, хищных сумчатых, и наконец, растительноядных, например кенгуру. В настоящее время кроме Австралии небольшое количество сумчатых имеется в Америке, тогда как ископаемые формы известны также и в Европе.
В Австралию попали также разные виды мышей и летучие мыши. Наконец вместе с человеком, который проник в Австралию из Азии, туда пришла и собака, которая затем одичала и известна как собака динго. Однако не только фауна млекопитающих Австралии так отлична и своеобразна в сравнении с фауной других материков. То же самое относится к птицам, а также многим другим группам животных и растений.
Своеобразие фауны Австралии можно объяснить лишь очень давней изоляцией ее от других материков. Благодаря этому эволюционный процесс мог происходить в другом направлении, чем на остальных материках, где на огромных территориях происходила миграция вновь возникших форм.
Так как географическая изоляция имеет такое огромное значения для эволюционных процессов, следует в свою очередь остановиться на различных видах изоляционных барьеров. Мы не можем забывать о том, что барьер, очень эффективный для одного вида, не представляет серьезного препятствия для распространения других видов. Для некоторых видов барьером могут служить высокие горные цепи, как, например, Гималаи в Азии, вершины которых покрыты вечным снегом. В этом случае барьером является не только высота над уровнем моря, но и резкие различия температуры.
Однако не для всех видов животных резкое изменение климата является барьером. Тигр может жить как в тропических джунглях, так и в районах покрытых снегом. Слоны также прекрасно переносят морозы, если только имеют достаточное количество воды. Ганнибал, выступая против Рима, имел большое количество слонов, вероятно африканских, с которыми переправился через Альпы. В настоящее время африканского слона можно встретить на очень большой высоте, в районе Килиманджаро, а индийские слоны живут как в жарких низменных районах Индии, так и на значительной высоте.
Гораздо более чувствительными к колебаниям температуры являются пресмыкающиеся. Известно, например, что крокодил может жить только в тропических или субтропических районах. Для некоторых животных недостаток влаги является решительной помехой для их распространения.
Таким барьером является, например, Сахара, которая до недавнего времени была залита морем, а в настоящее время служит преградой для миграции многих видов. Поэтому в эфиопскую область входит не вся Африка, а лишь часть ее к югу от Сахары.
Для других видов животных изоляционным барьером могут являться густые джунгли. Так например, когда в конце третичного периода обе Америки соединились, некоторые крупные млекопитающие, живущие в Северной Америке, например слоны и мастодонты, не могли проникнуть в Южную Америку через узкий Панамский перешеек, покрытый девственным лесом. Однако наиболее частым и наиболее эффективным для большинства животных является водный барьер, особенно соленые воды. Из позвоночных не могут преодолеть барьера соленой воды, земноводные, пресмыкающиеся и сухопутные млекопитающие, не считая летающих млекопитающих (летучие мыши); они могут преодолеть этот барьер в исключительных случаях. Земноводные чаще всего очень чувствительны к действию морской воды: в одинаковой степени как их личинки, так и взрослые формы. Лишь некоторые азиатские виды могут жить в солоноватой воде, а их головастики даже в соленой. Для млекопитающих, кроме морских, конечно, моря являются эффективным изоляционным барьером, если они не покрыты льдом. Хотя некоторые сухопутные млекопитающие прекрасно плавают, следует сомневаться в том, что они могли бы переплыть большие морские расстояния.
Для водных животных, как пресноводных, так и живущих в соленой воде, суша является изоляционным барьером. Кроме того, пресноводные животные чаще всего очень чувствительны к действию соленой воды, и наоборот, для морских животных пресная вода чаще всего является убийственной.
Несмотря, однако, на эти барьеры, в исключительных случаях может происходить распространение видов. Сухопутные животные иногда могут переплывать моря на уносимых стволах деревьев, ветвях или оторванных от суши так называемых плавающих островах. Это встречается не так редко, особенно в тропиках, где бушующие ураганы и ливни приводят к отрыву части суши вместе с находящимися там растениями и животными. Такие плавающие острова не раз встречались в сотнях миль от устий больших рек. Они обычно покрыты буйной растительностью и их иногда принимали за настоящие океанические острова. Убедились также в том, что такая большая змея, как боа, может на стволе дерева проплыть сотни морских миль. На далеком севере роль плавающих островов исполняют массы льда, на которых переплывают через Берингов пролив северные олени и белые медведи.
Летающие животные, как птицы, летучие мыши и насекомые, могут относительно легко преодолевать водные барьеры, если последние не очень широки. Но и при довольно больших расстояниях ветры помогают летающим животным преодолевать их. На ногах птиц могут быть перенесены разные виды низших животных, как, например, цисты одноклеточных, маленькие улитки и другие.
Одной из задач биогеографии является не только изучение различного рода изоляционных барьеров, но также и возможности преодоления их различными видами животных. Благодаря этим сведениям мы можем более подробно проанализировать все факторы, необходимые для того, чтобы понять распределение животных и растений, опираясь на принципе эволюции.
Необыкновенно важные доказательства из биогеографии в пользу эволюции приносит изучение фауны и флоры островов, как континентальных, так и океанических.
Континентальными островами называются острова, которые отделились от материков. Если континентальные острова возникли недавно, то отделяющее их от континента море неглубоко, а фауна и флора их очень сходны с фауной и флорой материнского континента. Так, например, геологи считают, что Британские Острова очень недавно отделились от Европы, примерно 7ССО лет назад. Это слишком короткий промежуток времени, чтобы изоляция могла привести к возникновению различия между европейскими видами и видами, живущими на Британских островах. Все виды животных и растений Британских островов идентичны европейским видам или родственны им.
Также и Японские Острова отделились от Азии недавно. Поэтому фауна Японских островов очень сходна с фауной соседней с ними части Азии. Если же континентальные острова отделились давно, то не только глубокие воды, отделяющие остров от суши, но и длительное время привело к возникновению значительного зоогеографического и фитогеографического различия.
Происхождение океанических островов является вулканическим, и в моменте возникновения они не имеют никаких представителей ни фауны, ни флоры континента. Однако позже на эти острова попадают некоторые представители растительного и животного мира с ближайшего материка, которые в свою очередь подвергаются дальнейшей эволюции. Изучение фауны и флоры океанических островов доставляет одно из наиболее убедительных косвенных доказательств эволюции.
На океанических островах, находящихся далеко от материка, не встречается земноводных и млекопитающих, так как эти животные, как правило, не могут преодолеть довольно широкого барьера соленой воды. Если же человек завезет туда различных млекопитающих и земноводных, то они могут хорошо приспособиться к жизни на океанических островах. Отсутствие на океанических островах различных форм этих животных вызвано не какими-то неблагоприятными условиями, а невозможностью преодоления больших пространств морских вод.
Изучая фауну и флору океанических островов мы замечали, что она является родственной фауне и флоре ближайшего континента, так как существуют наибольшие шансы случайного попадания живых организмов лишь с наиболее близкого континента. Таким образом, фауна и флора Галапагосских островов родственна фауне и флоре Южной Америки, тогда как фауна и флора островов Зеленого Мыса родственны фауне и флоре Африки, то есть ближайшего материка. Если океанический остров лежит относительно близко к материку, то хорошо летающие животные имеют возможность заселения его и скрещивания с особями того же вида, которые уже раньше попали на данный остров. Ввиду этого в данном случае не существует изоляции между видами материка и острова, что противодействует возникновению на острове новых видов из форм, которые туда проникли.
Если же остров лежит далеко от материка, то только совершенно случайно туда могут попасть даже хорошо летающие животные. Особи, которые оказались на таком острове, хорошо изолированы от материнских форм и не могут с ними скрещиваться. В этих условиях из первичного вида, прибывшего с ближайшего континента, иногда возникают новые виды, которые встречаются исключительно на этом острове, и которые называются эндемичными видами или эндемиками.
Ясно, что виды, не обладающие органами, служащими для полета, случайно попавшие на океанический остров, даже лежащий вблизи материка, могут через соответственно длительный промежуток времени образовать эндемичные виды. Чем более старым является океанический остров, тем больше были шансы попадания на него многих различных видов и тем более будут они отличаться от материнских форм. Иногда различия между материнскими формами и формами океанических островов имеют характер эндемичных подвидов, через более длительные промежутки времени возникают эндемичные виды и даже более высокие систематические категории, как роды и семейства.
Добржанский, (Dobzhansky) цитируя Циммермана, пишет о том, как ничтожны шансы проникновения видов животных на океанические острова, лежащие далеко от материков, и акклиматизация иммигрантов. На Гавайских островах можно встретить 3700 видов эндемичных насекомых и около 1000 эндемичных видов улиток. Циммерман доказывает, что эти 3700 видов насекомых развились из 250 видов, которые случайно попали на острова, а эндемичные виды улиток - из 25 видов иммигрантов. Можно рассчитать, что новый вид насекомых попадал на остров в среднем каждые 20000 лет, а улиток - 200000 лет.
Все перечисленные факты, а именно: отсутствие некоторых видов животных на океанических островах, родство фауны и флоры этих островов с фауной и флорой ближайшего материка, и, наконец, наличие эндемичных видов можно объяснить только путем постепенной эволюции растительного и животного мира.
Ниже приведем несколько избранных примеров специфичности фауны некоторых океанических островов. Галапагосские острова, известные со времен путешествия Дарвина, лежат на экваторе на расстоянии около 700 морских миль от западных берегов Южной Америки. 10 из 15 островов более крупных размеров это вулканические острова, окруженные глубокими водами. Здесь совершенно нет пресноводных рыб и земноводных. Из пресмыкающихся имеются огромные сухопутные черепахи, относящиеся к одному эндемичному виду, в состав которого входит больше десяти эндемичных подвидов, два эндемичных вида крупных ящериц (игуаны) и эндемичные змеи. Родственные виды ящериц и змей живут в Южной Америке.
На Галапагосских островах имеется также большое количество эндемических птиц, а между ними знаменитые вьюрки, образующие отдельное подсемейство (Geospizinae), которое включает 12 видов, относящихся к нескольким родам. Эти птицы развились на Галапагосских Островах из какого-то южноамериканского вида, приспосабливаясь к различным видам пищи, что наложило свою печать на формировании клюва у разных видов (рис. 28). Из млекопитающих животных встречаются только мелкие грызуны и один вид летучих мышей.

Рис. 28. Вьюрки с Галапагосского архипелага; по S.A. Barnett
Вся эндемичная фауна этих островов развилась из небольшого числа видов, происходящих из Южной Америки, которые в новых благоприятных для них условиях, и свободных от конкуренции развились эволюционным путем в различных направлениях, как, например, приведенные выше вьюрки. Вероятно протопластами всей фауны являлся один вид черепах, несколько видов ящериц, один или два вида змей, один вид грызунов и один - летучих мышей. Почти все виды насекомых являются эндемиками, относящимися к разным, и тоже эндемичным родам. То же самое относится к моллюскам.
Из высших растений примерно половина видов является эндемическими. Интересным является тот факт, что на Галапагосских островах имеется больше эндемичных форм, чем, например, на Азорских или Бермудских, хотя эти последние лежат дальше от ближайшего материка, чем Галапагосские. Однако этот факт легко можно объяснить. Галапагосские острова лежат в районе, где нет резких ветров, бушующих на северном Атлантике, которые облегчают перенесение организмов с суши на очень отдаленные острова.
Остров Святой Елены, на котором Наполеон I жил в британской неволе, лежит среди океана, на расстоянии 1100 миль от Африки и 1800 от Южной
Америки. Когда этот остров был впервые открыт, он был покрыт густыми девственными лесами, которые были уничтожены в XVI, XVII и XVIII веках. Таким образом, вместе с лесами было уничтожено большинство представителей фауны, которая в настоящее время была бы неоценимым сокровищем науки. Эта фауна была памятником древней фауны мезозойской эры, как бы музеем древних форм животных, лишь немногочисленные остатки которых сохранились до настоящего времени. На острове Святой Елены совершенно нет эндемичных млекопитающих, пресмыкающихся, пресноводных рыб, сухопутных птиц. Зато буйно представлен мир насекомых. Насекомые относятся почти исключительно к эндемичным видам и родам, а некоторые формы не имеют родственников в других местностях.
На Гавайях из сухопутных позвоночных живут два эндемичных вида ящериц, из которых один относится к эндемическому роду. Из водоплавающих птиц встречаем 24 вида, из которых 5 являются эндемичными. Даже одно из семейств птиц, включающее несколько родов, является эндемичным.
Таких примеров можно было бы привести гораздо больше. Объяснить их можно только благодаря принципу эволюции. Если отбросить эволюционную теорию, то факты биогеографии, становятся совершенно непонятными и хаотическими. Работа биогеографов еще не окончена. Эволюционная биогеография, датирующаяся от времен Дарвина, и, особенно, Уоллеса и других, имеет еще много проблем, которые следует решить. В этой работе неоценимую помощь оказывают результаты палеонтологических исследований. Приведем один пример.
Большой трудностью для биогеографов представляло объяснение особого размещения некоторых животных и растений. Так, например, тапиры живут только в юго-восточной Азии и центральной и Южной Америке. Араукарии растут в Австралии и Южной Америке, их совершенно нет в Африке и на северных материках. Некоторые биогеографы, стремясь объяснить это странное размещение некоторых форм животных и растений, предполагали, что современные материки в далеком прошлом были соединены между собой сушей. Считалось, что в давних геологических эпохах существовала сухопутная связь между Австралией, Южной Америкой и Африкой.
Однако не существует доказательств этой теории, а факты, приведенные выше, можно объяснить другим путем, принимая во внимание давнее распространение некоторых животных и растений. Так, например, тигры жили когда-то только в Северной Америке, Европе и Азии. Однако со временем они вымерли, оставаясь лишь в двух наиболее выдвинутых на юг пунктах их первоначального места пребывания. Также и араукарии когда-то росли в северном полушарии, о чем свидетельствуют, например, их окаменелые остатки в Аризоне. По мнению американского палеонтолога Симпсона, Антарктика никогда не была непосредственно соединена с южными частями других материков.