
Особенности природы умеренных пространств территории СССР
Они лучше известны для внеледниковых районов, т.к. здесь полнее, чем в ледниковых, сохранились отложения. Особенно это относится к бассейну Понто-Каспия, югу Западной Сибири и Дальнего Востока. В более северных районах верхнеплиоценовых отложений почти нет. Материалы по флоре, фауне и климату свидетельствуют, что во второй половине плиоцена природные условия постепенно приближались к современным.
Европейская часть СССР
На юге Европейской части была весьма своеобразная обстановка, обусловленная трансгрессиями и регрессиями Черного и Каспийского морей. Особенно значительными были колебания уровня и площади Каспия. Самой крупной была акчагыльская трансгрессия (рис. 6), распространявшаяся до бассейна Камы и низовьев Амударьи. На Кавказе береговая линия располагалась недалеко от Тбилиси. Крупной была и апшеронская трансгрессия. Трансгрессии Черного моря (таманская и более поздняя гурийская) были небольшими. Мощность акчагыльских и апшеронских отложений достигает местами более 3 км (например, в Куринской впадине).

Гиппарион и Этрусский носорог
Палеомагнитные данные указывают, что эти отложения формировались в эпохи Гаусса и Матуямы, а перекрывающие их бакинские отложения - уже в эпоху Брюнеса, к которой относится весь плейстоцен. В абсолютном летосчислении акчагыльские и таманские отложения формировались 1,7-3,5 млн. лет назад. Время формирования апшеронских и гурийских отложений ограничивается интервалом от 1,7 до 0,7-0,9 млн. лет назад.
Фауна моллюсков, остракод и фораминифер свидетельствует, что в различные фазы трансгрессий соленость моря заметно колебалась. Во время максимума она была ниже океанической. Морская фауна отличалась своеобразием. О происхождении акчагыльской фауны нет единства Мнения. Одними она считается сформировавшейся в процессе эволюции миоценовой фауны (Н.И. Андрусов и др.). Другими допускается ее образование в результате миграции океанической фауны. Скорее всего апшеронская фауна морских моллюсков формировалась из акчагыльской фауны и из мигрантов из Черного моря. Так, в таманских слоях черноморского бассейна, синхронизируемых с акчагылом, имеются типичные акчагыльские виды (Avimactra subcaspia, Cardium dombra и др.).
В последнее время начали появляться первые сведения о температурах вод палео-Каспия, определенные по химическому составу морских организмов. Так, К.М. Султанов и С.А. Исаев (1974) для акчагыльского бассейна в пределах Азербайджана указывают очень низкие температуры (13-14°). Несколько выше даются ими температуры для апшеронского века (17-18°). Они значительно (на 10-12°) ниже температур вод более древних бассейнов и немного ниже температур воды плейстоценовых бассейнов Каспия. В эпохи с низкими температурами резко сокращается накопление биогенных карбонатов (особенно в акчагыльское время).
На юге Русской равнины происходило формирование континентальных отложений аллювиального, дельтового, озерного и субаэрального (процессы, происходящие на суше) генезиса, образующих несколько самостоятельных стратиграфических горизонтов. К одному из них относятся хопровские отложения, распространенные в Приазовье и приуроченные к IX (40-50 м) террасе Дона. Это пески, глины и суглинки. В них известны костные остатки млекопитающих хопровского фаунистического комплекса. Более молодые ногайские отложения слагают более низкую террасу.
Аллювий хопровской и ногайской террас обычно перекрывается своеобразной толщей, известной в литературе под названием скифских или красно-бурых глин. Они распространены на огромной территории юга СССР: Молдавии, Украины, Приазовья, Приуралья и Зауралья, юге Западной и Восточной Сибири, Средней Азии, а также и за рубежом (в Монголии, Китае, Индии).
Встречаются они как на высоких террасах, так и на междуречьях, достигая 50-70 м мощности. Часто в них имеются мощные погребенные почвы (до 3-4 и более), а иногда и лессовые отложения. Скифские глины, судя по всему, формировались в переменно-влажном климате, сходном с климатом современного Средиземноморья. Переслаивание глин и почв-свидетельство частых Изменений природной обстановки. Скифские глины имеют примерно тот же возраст, что и апшеронские и акчагыльские отложения.

Трогонтерий и Саблезубый тигр
Палеомагнитные данные по хопровским, ногайским и скифским отложениям указывают на формирование их в эпохи Гауса и Матуямы. Хопровские отложения имеют калий-аргоновые датировки в 2,7-1,7 млн. лет. Ногайские отложения и самые молодые горизонты скифских глин датируются в 1,7-0,9 млн. лет. Более молодые отложения, с костными остатками животных тираспольского комплекса, относятся уже к эпохе Брюнеса.
Палеоботанические материалы свидетельствуют, что в акчагыле и апшероне были неоднократные перестройки растительности (Е.Н. Ананова, В.К. Немкова и др.). Однако крупнейшее изменение растительного покрова отмечено намного раньше, на границе миоцена и плиоцена, когда широко распространились степи. В это же время происходили и крупные изменения фауны млекопитающих. В акчагыле в бассейне нижней Камы, Среднем Поволжье, в Северном Прикаспии, т.е. южнее, чем ныне, была растительность таежного типа. В спорово-пыльцевых спектрах господствует пыльца ели с примесью пыльцы сосны, тсуги, пихты. Встречается и пыльца ряда экзотических видов липы, вяза, граба, карий, птерокарии и др. (рис. 7). Однако в сравнении с более ранними эпохами леса акчагыльского времени были беднее. Это следствие нарастающего похолодания. Оно продолжалось и в апшероне, когда в этих районах также господствовали леса, а южнее простиралась лесостепь и степь, которые напоминали современную саванну.
Распространение лесов в акчагыле и апшероне южнее, чем ныне, и наличие саванновых ландшафтов свидетельствуют о большей влажности климата. Это был результат изменения очертаний суши и моря, а также влияния трансгрессий Понто-Каспия.В. П. Гричук пишет, что для флоры того времени типично постепенное выпадение теплолюбивых экзотов - элементов восточноазиатской и североамериканской флоры - и увеличение роли представителей европейской флоры. Это - свидетельство похолодания климата.
Работы последних лет П.И. Дорофеева, В.К. Немковой и др. доказывают, что в акчагыле и апшероне происходило похолодание. Его результатом было глубокое проникновение на юг, вплоть до Башкирского Предуралья, Белоруссии, ряда холодолюбивых растений, например кустарниковой березы. Так, В.К. Немкова указывает, что в конце акчагыла в Башкирском Предуралье были холодные степи с березово-хвойным редколесьем. Флора апшерона, хотя и содержала некоторые теплолюбивые растения, тем не менее была
Данные по фауне млекопитающих, основные местонахождения которой находятся на юге, позволяют реконструировать палео - ландшафты и особенности эволюционного развития фауны. Главная заслуга в ее изучении, несомненно, принадлежит В.И. Громову (1948), выделившему несколько разновозрастных фаунистических комплексов. Верхнеплиоценовая фауна юга Русской равнины - родоначальница плейстоценовой и современной фауны этой территории.

Лошадь Стенона и Сиватерий
Для верхнего плиоцена характерны три фаунистических комплекса. Самый древний - молдавский комплекс. В его состав входили животные, типичные еще для гиппарионовой фауны: гиппарионы (трехпалые лошади), мастодонты, жирафы и др. Однако основное ядро было представлено новыми группами животных; слоны, носороги, быки, лошади и др., ставшими впоследствии типичными представителями плейстоценовой фауны. Особенно характерными животными были мастодонты (борзона и овернский) (рис. 7), носороги рода Dicerorhinus, лошади (Allohippus robustrus, A. stenonis). Вместе с ними обитали и гиппарионы (Hipparion cf. crassum). Более молодые комплексы (хопровский и таманский) в основном представлены теми же группами животных, но многие из них относились к новым родам и видам, стоящим на более высокой ступени развития. Вместе с тем отмечается и вымирание ряда животных: тапиров, динотериев, жирафов и др. или сокращение ареалов (мастодонты, гиппарионы и др.). Для хопровского комплекса типичен слон Громова, южный слон, этрусский носорог, лошади, эласмотерии и др. В последнее время Л.И. Алексеева (1975) хопровский комплекс подразделяет на раннюю и позднюю стадии. Для ранней стадии характерен слон Громова, а для поздней - типичная форма южного слона. В таманском комплексе обитала более поздняя форма южного слона. Лошади Стенона замещаются зюссенборнскими лошадьми. Появляются новые виды эласмотерия, зубров и др. Некоторые группы животных встречаются заметно реже и сокращают свои ареалы (мастодонты, саблезубые тигры, гиппарионы).
Каждый из древних комплексов отличается от молодых большим распространением древних групп животных. В молодых комплексах их место занимают " прогрессивные" формы, обитавшие в условиях более прохладного климата. Представители всех трех комплексов млекопитающих обитали в степных, лесостепных и саванновых ландшафтах. Изменения видового состава свидетельствуют об изменениях климата и ландшафтов. Климат становился все более прохладным, а растительность все более холодостойкой и жесткой, что отчетливо фиксируется в эволюции зубного аппарата хоботных и лошадиных. Однако фауна млекопитающих, так же как и флора, свидетельствует еще о вполне благоприятном климате.
Азиатская часть СССР
И здесь в верхнем плиоцене формировались разнообразные отложения. На юге Западной Сибири особенно широко распространены озерные, аллювиальные и субаэральные отложения кочковской свиты, достигающие местами 100-120 м мощности. По времени формирования они соответствуют акчагылу и апшерону. Из кочковской свиты получены интересные палеоботанические данные.В. П. Никитин для низов свиты выделяет флору барнаульского типа, весьма близкую к плейстоценовой. Растения, типичные для современной флоры юга Западной Сибири, достигают 60-65%, но встречаются и миоценовые теплолюбивые реликты (до 10-15%).В. П. Никитин считает, что в позднем плиоцене на юге Западной Сибири господствовали степи, а на севере Барабы - лесостепи. Таким образом, зональность здесь была близка к современной, хотя климат был благоприятнее. В более поздние фазы формирования кочковской свиты теплолюбивых растений становилось все меньше. Таким образом, флора современного типа и в Западной Сибири своими " корнями" уходит в " глубокий" плиоцен. Однако близкий к современному облик флора Западной Сибири приобрела раньше, чем флора Русской равнины.
Из кочковской свиты и ее аналогов известно много находок костных остатков млекопитающих (Э.А. Вангенгейм, В.С. Зажигин, А.Н. Мотузко и др.), относимых к нескольким самостоятельным комплексам, в общем соответствующих молдавскому, хапровскому и таманскому комплексам. Здесь также обитали мастодонты, слоны (Громова и южный), древние формы лошадей, носороги, эласмотерии, верблюды рода Paracamelus и др. Довольно многочисленны и мелкие млекопитающие (Mimomys pliocatnicus, М. praehungaricus, Pliomys sp. ochotona, Trogontherium minus и др.). Моллюски из кочковских отложений представлены большим числом видов. Многие из них обитают и ныне на юге Западной Сибири, но имеются и теплолюбивые вымершие виды (Vertigo uralica. Gastrocopta huttoniana и др.).
В Прибайкалье и Забайкалье шло формирование субаэральных красноцветных карбонатных отложений чикойской свиты (галечники, пески, глины, прослои торфа). В районах наибольших прогибаний, например в Тункинской впадине, Э.И. Равский (1972) указывает 500-метровые толщи отложений этого возраста. Во время формирования чикойской свиты в этом районе сначала произрастали хвойные леса с примесью широколиственных пород, которые позже сменились лиственными лесами. В этих формациях постепенно вымирают наиболее теплолюбивые растения, их место занимают виды, приспособленные к менее благоприятному климату.
Однако данные спорово-пыльцевого анализа " противоречат" литологии чикойской свиты. Так, ее отложения красноцветны, что свидетельствует о формировании свиты в довольно жарких и временами сухих условиях. Скорее всего это противоречие возникло из-за того, что при реконструкциях использовалась переотложенная пыльца. Об этом же свидетельствуют и находки костных остатков млекопитающих чикойского комплекса (Вангенгейм, 1965). В его составе довольно древние формы в основном центральноазиатского происхождения: несколько видов гиппарионов, газели, носороги, цокоры, бобры, пищухи.
Каковы же особенности палеогеографии конца неогена юга Дальнего Востока? Современная природа этого района имеет много специфического: в здешней флоре обильны неогеновые и даже палеогеновые реликты субтропического происхождения, например пробковое дерево, маньчжурский орех, дикий виноград, лианы. Они свидетельствуют, что юг Дальнего Востока не претерпел сильных климатических изменений. Плиоценовые отложения здесь широко распространены, имеют большие мощности (сотни метров) и залегают нередко ниже уровня океана. Палинологические данные указывают, что в конце плиоцена на юге Дальнего Востока господствовали хвойно-широколиственные леса, в которых было много теплолюбивых элементов (особенно в Южном Приморье). Следовательно, климат был теплым и влажным. Вымирание самых теплолюбивых видов падает на конец плиоцена и отражает общепланетарное похолодание.
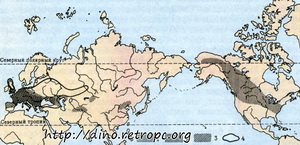
Рис. 7. Распространение мастодонтов, южного и трогонтериевого слонов: 1 - овернский мастодонт; 2 - американский мастодонт; 3 - южный слон; 4 - трогонтериевый слон;