
Сравнительно-эмбриологические доказательства эволюции
Постепенное формирование понятий, связанных с интерпретацией фактов, почерпнутых из сравнительной эмбриологии, является очень ярким примером из истории биологических наук и эволюционизма. Поэтому, перед тем, как перейти к изложению современных взглядов, представим коротко историю вопроса. Наша задача облегчается тем, что появились две статьи J. Oppenheimer и А.О. Lodejoy, в которых с большой проницательностью подчеркнуты основные черты воззрений старых биологов на факты из эмбриологии, касающиеся эволюции.
История теоретического исследования эмбриологических данных берет свое начало задолго бо того, как Дарвин начал свою научную деятельность и даже перед тем, как Ламарк опубликовал свою "Философию зоологии". Начиная с 1797 г., то есть от эпохи Гёте и Autenrieth и кончая Геккелем в 1866 г. семьдесят два автора обращали внимание на сходство эмбриологического развития различных форм, или на повторение стадий более низких форм эмбрионами более высоко развитых. В этом втором случае говорим о рекапитуляции.
Однако не подлежит сомнению, что в периоде, предшествовавшем Дарвину, наибольшее значение имеет научное творчество Карла Э. Бэра, который является истинным основоположником современной эмбриологии, как отдельной отрасли биологических наук, связывающей вопросы эмбриологии с вопросами эволюции. Как подчеркивает Оппенгеймер, Бэр, как каждая выдающаяся личность, был человеком необыкновенно сложным, и в свете этого следует рассматривать его отношение к принципам эволюции Дарвина. Несомненным является тот факт, что Бэр еще перед 1859 г. высказал мысли, которые можно интерпретировать в эволюционном смысле. И правдой является также тот факт, что после выступления Дарвина, Бэр стал противником эволюции и остался таким уже до конца жизни.
Основной труд Бэра носит заглавие: "О развитии животных. Наблюдения и размышления". Первая и вторая часть этого труда были опубликованы в 1828 и 1837 г., последние главы второй части вышли уже после смерти автора в 1888 г. Еще перед тем, как была издана первая часть книги, французский анатом Serres в 1824 г. отчетливо вводит понятие принципа рекапитуляции. "Человек становится человеком, проходя переходные организационные стадии, в которых вначале он похож на рыбу, затем на пресмыкающееся, птицу и млекопитающее".
Стоит отметить, что Serres своей концепции не связывал с какой бы то ни было эволюционной мыслью. В 1867 г. в заметке, представленной Французской Академии Наук, он пишет, что теория Ламарка, представленная снова Дарвином, противоречит научным фактам. Однако не это важно. Важным является то, что Бэр уже в первой части своего труда выступил против рекапитуляции, то есть повторения эмбрионами высших животных форм низших зрелых организмов.
Бэр отчетливо пишет, что "эмбрион высших форм животных никогда не похож на низшее животное, а лишь на зародыш этих форм". Автор, однако, не приписывает этого сходства зародышей какому-то родству, а считает, что при эмбриональном развитии вначале проявляются общие признаки, свойственные большей группе животных, а затем признаки все более специальные. Материал, однообразный в процессе развития, преобразуется в материал все более дифференцированный, гетерогенный. "Структура гетерогенная, специальная, развивается из более однородной, общей, путем постепенного изменения".
Хотя Бэр еще перед выступлением Дарвина высказывал воззрения, свидетельствовавшие о том, что определенные эволюционные концепции не были ему чужды, он не был эволюционистом и позже открыто выступал против теории Дарвина. Он никогда не пытался даже объяснить сходства зародышей разных животных с точки зрения эволюции, а лишь с точки зрения принятой им теории развития от однородной стадии к дифференцированной. В этом духе следует тоже интерпретировать его высказывание:
"Зародыш позвоночного уже с самого начала является позвоночным и никогда не проявляет сходства с беспозвоночным животным. Мы не знаем такого животного, которое бы относилось к позвоночным, а гистологически и морфологически было бы так мало дифференцировано, как эмбрионы позвоночных. Поэтому эмбрионы позвоночных животных в своем развитии никогда не претерпевают никаких изменений форм животных... Зародыши млекопитающих, птиц, ящериц и змей, а также, вероятно, черепах на ранних стадиях развития, необыкновенно похожи друг на друга как по внешнему виду, так и развитию отдельных органов, настолько, что эти зародыши можно различить только по их величине.
У меня самого хранятся два небольших эмбриона в спирту, которые я в свое время забыл снабдить этикетами, и я теперь не в состоянии определить к какому классу они относятся: это могут быть маленькие ящерицы, птицы или молоденькие млекопитающие, так они похожи друг на друга формой своей головы и туловища. Эти эмбрионы еще не имеют конечностей. А если даже и имели, то это тоже не помогло бы в определении вида эмбрионов, так как в первом периоде развития ноги ящериц и млекопитающих, крылья и ноги птиц, руки и ноги людей развиваются из той же основной формы".
Дарвин, который до конца жизни с трудом читал по-немецки, не знал в оригинале трудов Бэра и основывался только на переводе Гексли и работах Карпентера и Barry, в которых нашел описание результатов исследований Бэра. Дарвин, который в двух первых изданиях "Происхождения видов" ошибочно приписывал цитированное выше высказывание Бэра Агассицу, сразу оценил значение эмбриологических данных для теории эволюции. Для него сходство эмбрионального развития животных является доказательством общего происхождения этих форм, то есть имеет такое же значение, как результаты изучения сравнительной анатомии в отношении гомологических органов.
Однако Дарвин не остановился на этом. Несколько в первых набросках своей теории в 1842 и 1844 г. он останавливается только на сходстве зародышей, как доказательстве их общего родового происхождения, постольку уже в первом издании "Происхождения видов" пишет: "Ввиду того, что эмбриональная стадия каждого вида или группы видов показывает нам строение их менее измененных давних предков, мы отчетливо видим, почему давние и вымершие формы жизни должны быть сходны с зародышами их производных форм". Здесь Дарвин впервые говорит о рекапитуляции, мысль о которой была в последствии развита Ф. Мюллером и Геккелем и сформулирована в виде биогенетического закона.
По этому закону онтогенез является кратким повторением филогенеза, то есть индивидуальное развитие является повторением родового, эволюционного развития. Если мы подробно знаем отдельные этапы онтогенеза, то на этом основании, по закону Геккеля, мы можем восстановить очередные фазы филогенеза, познакомиться с очередными предками рассматриваемых форм, и представить в виде генеалогического дерева развитие данной группы животного или растительного мира. Кроме того, так как решающее влияние на онтогенез оказывает филогенез, вопросы эмбриологии ограничиваются исключительно эволюционными исследованиями, и нет необходимости доискиваться каких-то других факторов, управляющих онтогенетическим развитием.
Геккель, который был яростным поклонником такого воззрения, отмечал, однако, что несмотря ни на что, изучение онтогенеза не может дать нам полной картины филогенеза. Каждый зародыш в своем развитии, кроме проявления черт, являющихся точной рекапитуляцией, то есть палингенетических, обладает также признаками, являющимися проявлением приспособления зародыша к специфическим условиям эмбрионального развития, то есть ценогенетическими.
Зато такие признаки, как наличие жаберных дуг и щелей, которые имеют место в развитии пресмыкающихся, птиц и млекопитающих, не исключая человека, следует, по мнению Геккеля, считать признаками палингенетическими, отражающими те стадии развития предков, которые жили в воде и дышали жабрами. К палингенетическим признакам Геккель относит хорошо развитую в определенной стадии развития зародыша человека, хвостовую часть, как и оволосение всего тела зародыша.
Дарвин придавал большое значение эмбриологическим данным для обоснования эволюционной теории, и потому был несколько удивлен тем, что рецензенты первого издания "Происхождения видов" не обратили внимания именно на ту главу, в которой он излагал данные из эмбриологии. По мере появления новых изданий "Происхождения видов", Дарвин все больше использовал материалы и взгляды своих сторонников. В 1864 г. выступил Ф. Мюллер, и Дарвин в четвертом издании "Происхождения видов", базируясь на взглядах этого автора, считает, что "в очень отдаленные времена вероятно существовало зрелое животное, похожее на науплиуса (личинку ракообразных), которое в процессе дальнейшего развития дало начало разным группам ракообразных.
В 1866 г. была издана книга Геккеля "Общая морфология", в которой этот автор развивает дальше и расширяет теорию рекапитуляции. Яйцеклетка животных соответствует одноклеточным предкам многоклеточных животных. Эмбриональная стадия бластулы соответствует зрелой форме предка, которого Геккель называет бластея, тогда как двулистковая гаструла соответствует гипотетическому предку, названному гастрея.
Как подчеркивает де Бир, теория рекапитуляции Геккеля была принята современными ему эволюционистами без возражений. Некоторые из авторов, занимавшиеся исследованием окаменелостей, старались даже на своем материале подтвердить правильность предпосылок Геккеля. Однако вскоре результаты этих исследований были опровергнуты, а одновременно появились критические замечания по адресу теоретических предпосылок концепции Геккеля. Совершенно непонятно, каким образом могли быть включены в процесс эмбрионального развития стадии зрелых форм. Несмотря на все сходства, зародыши разных животных неидентичны, неодинаковыми являются и их яйцеклетки. Жаберные щели зародышей млекопитающих не являются рекапитуляцией рыбы, их можно только сравнивать с образованиями, имеющими место у эмбрионов рыб.
В связи с все более прогрессивным развитием генетики и разработкой Морганом и его сотрудниками хромосомной теории наследственности, ученые--генетики, изучающие вопросы эволюции, были вынуждены заняться также интерпретацией, в свете науки о наследственности, фактов, доставляемых сравнительной эмбриологией.
Первым из авторов, который рассматривал доказательства из эмбриологии с точки зрения современной генетики, был Морган. Морган решительно выступал против биогенетического закона. Оп считал, что материал, полученный при изучении сравнительной эмбриологии, может найти свое логическое объяснение только базируясь на эволюционных принципах.
"Факты говорят сами за себя. Это жаберные щели, обнаруженные у зародышей птиц и млекопитающих, дорзальная хорда у зародышей, уступающая позднее свое место позвоночнику, очередность изменений сосудистой системы, последовательное развитие выделительной системы у млекопитающих в виде предпочки, первичной и вторичной почки, история развития половых органов и другие. Эти и многие другие такие же факты находят, как мне кажется, достаточное объяснение в эволюционной теории, как отображение давней истории в эмбриональном развитии высших групп".
"Обратим внимание например, на позвоночных. Согласно теории эволюции, высшие группы происходят из видов, сходных с примитивными видами, которые жили в давние времена. Если эволюционные изменения происходят главным образом в конечных стадиях онтогенетического развития, то можно понять, почему соответствующие стадии развития высших и низших обладают сходствами".
"Жаберные щели являются хорошим примером. У зародышей костистых рыб жаберные щели развиваются на ранних этапах развития когда зародыш еще покрыт яйцевой оболочкой. Щели эти остаются, и в процессе дальнейшего развития становятся постоянными органами дыхания зрелых форм. У птиц и млекопитающих щели также развиваются на очень раннем этапе и сходным образом, как у зародышей рыб. Однако позже большинство щелей закрывается и исчезает. Из двух или трех щелей в дальнейшем развиваются иные органы"...
"Такое развитие можно связывать с теорией эволюции, если мы объясняем эти факты таким образом, что некоторые стадии зародыша рыбы сохранились без значительных изменений у ранних зародышей птиц и млекопитающих. Такая интерпретация не только согласована с нашим ощущением непрерывности и родства, но не содержит никаких трансцендентальных предпосылок, которые принимает теория рекапитуляции, переносящая зрелые формы предков на ранние этапы развития более поздних форм".
Данные сравнительной эмбриологии, кроме того, показали, что вопрос соответствующей интерпретации стадий развития является гораздо более сложным, чем предполагали первые эволюционисты. Особые заслуги в этом вопросе принадлежат А.Н. Северцову и его сотрудникам, а также Гарстангу (Garstang).
Гарстанг исследуя стадии развития разных ракообразных, установил, что организмы во время эмбрионального развития не рекапитулируют зрелых форм своих предков, а повторяют лишь до определенного момента соответствующие стадии эмбрионального развития протопластов. Кроме того Гарстанг установил, что многие личинки не являются повторением каких то форм давнего генеалогического дерева, а что они специально приспособлены к распространению в море или пресных водах. Обращено также внимание на то, что в истории филогенеза не только зрелые формы могли дать начало новым видам организмов, но и незрелые, юные формы. В этом случае мы говорим о так называемом педоморфозе.
Педоморфоз двояким путем может играть роль в эволюции. Во-первых, новое эволюционное изменение появляется у юной формы, вызывая изменения как у форм, так и у зрелых производных форм. Хорошим примером являются брюхоногие моллюски. Совершенно ясно, особенно после того, как в 1957 г.Н. Lemche открыл примитивную метамерию у симметричной Neopilina что первичные, древние моллюски были симметричными. Совершенно ясно также, на основании данных сравнительной анатомии и палеонтологии, что у первичных брюхоногих произошел оборот тела на 180°, что привело к ассиметрии всех их органов.
Рис. 25. Neopilina. Слева с брюшной стороны, справа - дорсальной; по L. Chopard
Трудно было представить себе, каким образом мог наступить этот оборот благодаря, постепенным, очередным изменениям, наступающим у зрелых форм. В эмбриональном периоде развития оборот происходит внезапно на ранних стадиях развития личинки. Оборот завершается в течение нескольких часов в результате отсутствия механической стабилизации личинки и сокращения мышц, развитых асимметрично по одной стороне, как это установил D.R. Croft в 1955 г.
Изменения во всем строении личинки происходят внезапно и вполне возможно, что также в истории рода изменения в строении произошли таким же образом на этапе личинки. В другом случае педоморфоза мы имеем дело с сохранением у зрелых форм отдельных признаков личинки. Приведем один пример, почерпнутый у Гарстанга. Автор этот предполагает, что личинка морской звезды, так называемая аурикулария, могла послужить переходным пунктом для всех хордовых, к которым относятся также позвоночные. Аурикулария во многом сходна с некоторыми хордовыми, скелет ее происходит из мезодермы, а мышцы иглокожих, к которым относится морская звезда, по своему биохимическому строению сходны с мышцами хордовых.
Никто, конечно, даже не предполагает, что хордовые могут развиться из зрелых форм иглокожих. Если же предположение Гарсганга является правильным, то в таком случае личиночная стадия путем педоформоза могла привести к эволюционному развитию всего огромного типа хордовых, включая и позвоночных животных.
Проблемы педоморфоза базируются не только на предположениях; известно, что один из видов хвостатых земноводных, живущих в Америке, сохраняет нормальные признаки личинки, не подвергается преобразованию и как личинка размножается. Это явление называется неотенией.
Несомненные доказательства, собранные учеными на поле сравнительной эмбриологии, имеют очень большое значение, не только подкрепляя сам принцип эволюции. В конкретных случаях, указывая на сходство между зародышами, они могут послужить познанию родовой истории. Как анатомическое сходство указывает на генетическое сходство, а этим и на родственную связь между организмами, так и эмбриологическое сходство является результатом определенной общности наследственного материала, то есть генов. НеСМОТРЯ на действие наследственной по П. Грассэ. изменчивости, (возникающей в результате мутации и рекомбинации генов в процессе полового развития), подвергающейся постоянному действию естественного отбора, неизбежно сохраняются определенные группировки генов или их систем, что является непосредственной причиной анатомического и эмбриологического сходства.
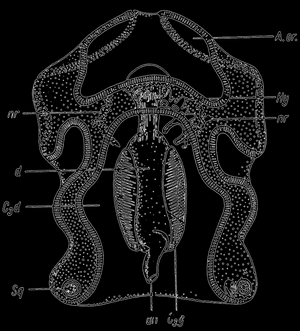
Рис. 26. Auricularia A.or - предротовая полость; С3d - полость тела; d - пищеварительный тракт; Ну - hydrocel, nr - нервная система; Sq - скелетные части;
Ясно, что эволюционные изменения могут касаться всех фаз онтогенеза, то есть могут приводить к изменениям не только зрелых организмов, но также и эмбрионов, даже на первых этапах развития. Тем не менее, однако, признаки, характеризующие ранние фазы развития, должны отличаться большим консерватизмом, чем более поздние, так как изменения на более ранних этапах развития в свою очередь должны привести к большим изменениям в процессе дальнейшего развития. Изменение, например, типа дробления вызовет изменения в процессе гаструляции, как и всех следующих стадиях. Поэтому наследственные изменения, проявляющиеся на ранних этапах, гораздо чаще приводят к летальным изменениям, чем таковые в более поздние периоды онтогенеза. Отсюда ранние стадии являются более консервативными и поэтому, благодаря своему сходству, могут служить нам информациями относительно эволюционного родства.
Hill и де Бир установили, например, что у зародышей сумчатых появляется зачаток так называемого зуба, который яйцеродным предкам служил для разбивания скорлупы яйца. Известно, что живородящие сумчатые появились примерно 75 миллионов лет назад. А однако зачаток древнего зародышевого органа сохранился до сегодняшнего дня.
Де Бир, разбирая в связи с юбилеем значение трудов Дарвина для эволюционно-эмбриологических исследований, подчеркивает, что "современные взгляды на взаимоотношение эмбриологии и эволюции в основном соответствуют тем взглядам, к которым Дарвин пришел в своем труде в 1844 г., и которые с многочисленными примечаниями были помещены в "Происхождении видов". Если мы выбросим из этого труда то, что Дарвин цитирует за Агассицом и Мюллером и что он сам, no-существу, принимал с оговоркой, то легко убедимся, как объективно обоснованными были его выводы относительно значения эмбриологии для эволюции"...
***
С того времени, как теория наследственности нашла свое обоснование в хромосомной генетике, большое значение имеет изучение комплексов хромосом, так как это проливает дополнительный свет на теорию эволюции. Так, например, как у пресмыкающихся, так и у птиц, в комплексах хромосом кроме крупных хромосом встречаются также и мелкие, называемые микросомами, что указывает на родство этих двух классов.
Интересно, что у клоачных, то есть у утконоса и ехидны, также имеются микросомы, тогда, как у других млекопитающих они отсутствуют. Отмечается также значительная разница между комплексами хромосом сумчатых и плацентных млекопитающих. Сумчатые отличаются небольшим количеством хромосом, тогда как плацентарные имеют большое количество более мелких хромосом. Кариология, то есть наука о комплексах хромосом, может оказаться полезной в систематике, помогая определить происхождение видов. Оказалось, например, что два вида ежей, живущих в Европе, совершенно не отличаются комплексом своих хромосом, и поэтому их следует считать подвидами (Иордан).
Основываясь на кариологических исследованиях редкого вида грызуна лесной мышовки - Sicista betulina - следует этот вид отнести к отдельному подсемейству (Валькновска). Домашняя собака, как и волк, имеет 78 хромосом, тогда, как шакал - 74. Исходя из количества хромосом и их структуры Маттей (Matthey) считает, что в эволюции собаки шакал не принимал никакого участия. Если бы собака произошла в результате скрещивания волка с шакалом, то в ее наборе хромосом должны были сохраниться также хромосомы шакала.