
Микроэволюция
В одной из предыдущих глав мы останавливались на выступлениях Дженкина и Кельвина против теории, провозглашенной Дарвиным. После их выступления Дарвин старался использовать все данные, свидетельствующие с одной стороны о действии отбора в природе, с другой - доказывающие быстрый темп эволюционных изменений. Поэтому в своем труде "О происхождении человека" он цитирует сообщение, происходящее от одного охотника из Соединенных Штатов, что в определенной местности олени с неразветвленными рогами встречаются все чаще. Мнимое эволюционное изменение, происходящее на протяжении короткого срока, оказалось иллюзией. В этом случае речь шла о годовалых оленях, у которых рога еще не ветвятся.
В настоящее время мы однако хорошо знаем, что на глазах у Дарвина в Великобритании происходил процесс, как бы образно представляющий течение эволюции. Однако его не заметил ни сам Дарвин, создатель теории естественного отбора, ни кто бы то не было из эволюционистов в течение долгих десятилетий. Поводом, возможно, было то, что продолжатели Дарвина направляли свои усилия главным образом не на доказательства правильности теории отбора, а на собирание все большего количества доказательств самого факта эволюции.
В данной главе мы постараемся кратко представить результаты исследований, обнаруживающих эволюционные изменения, происходящие в природе, которые можно изучать при помощи генетики и экологии. При помощи этих наук можно лучше всего проследить за эволюционными изменениями в масштабе, доступном нашему непосредственному наблюдению.
Эволюционный процесс, приводящий к образованию новых видов или более высоких систематических единиц, является процессом столь длительным, что его невозможно проследить. Мы можем судить о нем, лишь на основании косвенных доказательств или палеонтологических данных. Но если мы не можем проследить непосредственно макроэволюционных изменений, изменений в большом масштабе, то мы можем с успехом следить за так называемыми микроэволюционными процессами, то есть мелкими изменениями, заметными в популяциях. Особенно последние десятилетия доставили науке непосредственные доказательства микроэволюционных процессов, происходящих в природе. На них следует остановиться более подробно.
Условия, существующие на территории Великобритании значительно изменились от начала промышленной революции, то есть от второй половины XVIII века. От этого времени в больших промышленных районах начали выпадать во все большем количестве частички сажи, покрывающие стволы и ветви деревьев, листья и поверхность почвы. Кроме того налет сажи привел к гибели лишайников, покрывающих кору деревьев, а в районах с развитой промышленностью и утесы покрылись сажей.
Изменившиеся таким образом условия привели не только к гибели лишайников, то есть организмов, построенных по существу из двух разных растений, а именно грибов и водорослей. Одновременно изменилась окраска многих видов ночных бабочек, которые проводят день, сидя неподвижно на коре деревьев. В одной лишь Англии было отмечено, что больше, чем 70 видов ночных бабочек в быстром темпе изменило свою окраску из светлой на темную. Аналогичные изменения обнаружены в фауне ночных бабочек в промышленных районах многих стран Европы, как Франция, Германия, Польша, Чехословакия, а также в Канаде и Соединенных Штатах. Темных, меланических форм никогда не удается обнаружить в тропических странах.
Первый образец меланической ночной бабочки, Biston betularia, был пойман в Манчестере в 1848 г. Меланические формы, вначале очень редкие, начали распространяться в промышленных районах, вытесняя старые, типичные белые формы. В 1953г. в Бирмингаме 90% составляли меланические формы, и только 10% - светлые.
Это явление английский биолог Heslop Harrison пытался объяснить следующим образом. Он считал, что промышленный меланизм вызван специальной субстанцией, содержащейся в фабричном дыму. По его мнению здесь играют роль сернокислый магний и азотнокислый свинец. Он считал, что благодаря тому, что личинки съедают листья, покрытые сажей, большое количество бабочек, развивающихся из этих личинок, обладает темной окраской и что этот признак в дальнейшем наследуется согласно закону Менделя.
Работа Heslop Harrison, опубликованная в 1926 г., вызвала большой интерес, однако способ, в который автор объяснял данное явление, не был встречен с живым откликом. Уже прошло время, когда принимались концепции, основанные на принципах ламаркизма, провозглашающего наследование приобретенных признаков. Правда, в данном случае в игру могли входить явления мутации, вызванной действием химических факторов, содержащихся в саже и являющихся мутагенами. Однако, как вытекает из данных
Harrison, частота этих мутаций превышала бы все, что мы знаем о действии мутагенов, так как равнялась бы 8%.
Лишь работы оксфордских ученых позволили произвести правильный анализ явления промышленного меланизма. Стоит напомнить, что пионером изучения микроэволюционных изменений в природе был Е.В. Ford, который начал свою работу еще перед первой мировой войной. В настоящее время группа исследователей, ведущих свою работу в генетических лабораториях Оксфордского Университета, занимается одновременно экологическими и генетическими исканиями. Они создают базу для развития новой ветви генетики, то есть эволюционной генетики, целью которой явится прежде всего изучение микроэволюционных процессов, происходящих в природе, и контроль экспериментальных данных, полученных в лабораториях Университета.

Рис. 56. Промышленный меланизм у Biston betularia: a - меланистическая и почти невидимая типичная форма (светлая) на светлом стволе, b - на темном стволе почти невидима меланическая форма; по S.A. BarnettBiston betularia: a - меланистическая и почти невидимая типичная форма (светлая) на светлом стволе, b - на темном стволе почти невидима меланическая форма; по S.A. Barnett">
Изучение явления промышленного меланизма производилось главным образом на Biston betularia. Зрелая форма этой ночной бабочки появляется от мая до августа. Бабочки летают только ночью, а днем сидят на коре деревьев. Из отложенных яичек развиваются личинки, то есть гусеницы, питающиеся листьями. Они появляются начиная от июня. Осенью гусеницы превращаются в куколки и весной в мае из куколок вылупляются зрелые формы - бабочки. Светлые формы которые имеют крылья белого цвета с темными неправильной формы полосками и пятнышками, сидя поперек или косо на коре деревьев, покрытых лишайниками и непокрытых сажей, совершенно не видны. Их окраска является покровительственной.
Доказательством сказанного является фотография белой формы, сидящей на дереве, из сельскохозяйственных районов Англии, где встречаются почти исключительно белые формы (рис. 56). Зато, как это хорошо видно на фотографиях, светлые экземпляры хорошо видны на деревьях промышленных районов. Дело с меланистической формой обстоит как раз наоборот. Ее почти не видно на деревьях, покрытых сажей, зато она бросается в глаза на деревьях без сажи, покрытых лишайниками.
Из вышесказанного можно было бы предположить, что меланистические формы, появляющиеся в сельскохозяйственных районах, хуже приспособлены к условиям и уничтожаются птицами, охотящимися днем, в гораздо большем количестве, чем светлые формы. Естественный отбор, таким образом, элиминирует появляющиеся время от времени меланистические формы, как хуже приспособленные к жизни в данных конкретных условиях. Иначе действует отбор в промышленных районах, где лучше приспособленными являются меланистические формы.
Однако это предположение следовало проверить в двух направлениях, экологическом и генетическом. Экологическими исследованиями занимался главным образом Kettlewell. Бабочек отмечали цветными точками на нижней поверхности крыльев, применяя ежедневно краску другого цвета, и выпускали их на свободу. Затем вылавливали бабочек, привлекаемых светом. Оказалось, что если в районах, не покрытых сажей, выпускали светлых и темных бабочек, то позже вылавливали гораздо больше светлых форм. И наоборот, в промышленных районах вылавливали больше меланистических меченых форм, чем светлых. Оказывается, что в зависимости от окраски и условий среды, выживает больше особей, приспособленных своей окраской к среде.
Данные Kettlewell встретились, однако, с многочисленными возражениями как со стороны орнитологов, так и энтомологов, которые считали, что днем птицы никогда не ловят ночных бабочек, сидящих неподвижно на коре деревьев. В то же время Kettlewell и его сотрудники обнаружили, что птицы из семейств славек и синиц действительно ловят очень быстро сидящих на коре деревьев ночных бабочек, причем в сельскохозяйственных районах быстрее вылавливают темные формы, а в промышленных - светлые. Эти наблюдения затем были документированы фильмом, заснятым Тинбергеном (Tinbergen).
На приведенной карте (рис. 57) мы видим четкую корреляцию между заселением сельскохозяйственных районов светлыми формами и промышленных - меланистическими. Однако белые формы все реже, в сравнении с темными, встречаются в восточной Англии, хотя там и нет крупных промышленных районов.
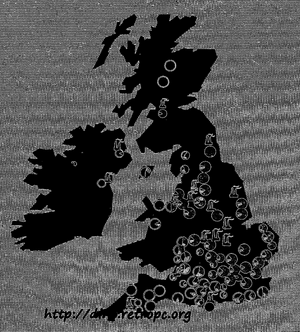
Рис. 57. Размещение светлых и темных форм Biston betularia на территории Великобритании. Чёрная часть круга обозначает количество тёмных форм в популяции В. betularia зачерченная часть - количество тёмных форм другого вида, insularia; белая часть круга соответствует количеству светлых форм. - Обозначены также промышленные центры, из которых западные ветры переносят сажу на восточные, сельскохозяйственные районы
Это легко объяснить. Благодаря тому, что ветры на юге Англии обычно дуют в направлении с запада на восток, сельскохозяйственные районы на востоке покрыты обильным количеством сажи, перенесенной ветром в сельскохозяйственные районы. Несмотря на отсутствие крупных промышленных предприятий в этих районах, меланистические формы там лучше приспособлены, чем светлые.
В дальнейшем генетические исследования показали, что меланистические формы являются результатом действия отдельного гена. Обозначим ген светлой окраски буквой с. Гомозигота ее будет отличаться светлой окраской. Доминирующим аллелем этого гена является доминирующий ген С. Светлые формы называются типичными, темные - саrbопаria. Ввиду того, что ген С является доминирующим, то как гомозигота СС, так и гетерозигота Сс имеют темную окраску.
Затем обнаружили, что ген С вызывает не только изменение окраски, а влияет также и на другие признаки. Гусеницы формы carbonaria являются более стойкими. Замечены также некоторые характерные различия в поведении типичной и меланической форм. В холодные ночи самцов типичной, светлой формы манят больше светлые самки, чем темные. Во время теплых ночей темные самки больше привлекают светлых самцов. Кроме того обе формы отличаются еще некоторыми признаками, как физиологическими, так и темпом размножения.

Рис. 58. Различные формы Bistort betularia, a - типичная светлая форма, b) форма insularia с, d, - формы carbonaria, причем форма (d) является экспонатом, пойманным в XIX в. Она более светлая, чем современная меланическая форма (с). Среди меланических форм окраска типа insularia зависит от наличия другого гена, чем окраска типа carbonaria; по P.M. Sheppardinsularia с, d, - формы carbonaria, причем форма (d) является экспонатом, пойманным в XIX в. Она более светлая, чем современная меланическая форма (с). Среди меланических форм окраска типа insularia зависит от наличия другого гена, чем окраска типа carbonaria; по P.M. Sheppard">
Kettlewell заметил еще одно очень интересное различие в поведении типичной формы и carbonaria. Оно касается подыскивания соответствующего цвета фона. Хотя наблюдения еще не закончены, результаты, полученные до настоящего времени, указывают на то, что типичные формы садятся на светлые стволы, a carbonaria - на темные. Этот факт Kettlewell объясняет принципом контрастной оценки.
"Этот механизм основан на том, что одна часть глаза бабочки воспринимает цвет субстрата, тогда как другая - собственный цвет. Таким образом, сравниваются обе окраски. Если они соответствуют друг другу, то бабочка остается на месте, если же они отличаются друг от друга, то возникает контрастный конфликт, бабочка улетает. Наблюдения показали, что бабочки не успокаиваются до тех пор, пока существует конфликт окраски".
Таким образом, мы видим, что изменению окраски сопутствует ряд других изменений, касающихся физиологических свойств, развития и поведения. В этом случае мы имеем дело с плейотропным действием гена на цвет, или же разные свойства зависят от разных генов, которые, однако, так сильно связаны между собой, что замена между ними (crossing over) может произойти лишь в виде исключения.
Мы уже отмечали, что ген С является доминирующим геном, так что фенотипически не существует разницы между гомозиготами СС и гетерозиготами Сс. Однако замечено, что первые пойманные представители формы carbonaria, относящиеся к половине прошлого века, были более светлыми, чем современные. Вероятно раньше ген С не обладал полным доминированием в отношении к аллели с.
Предположим, что мы наблюдаем сельскохозяйственный район, не загрязненный фабричными осадками. Если в этих условиях появится меланистическая мутация, то этот ген не имеет никаких шансов распространения, так как особи темного цвета вылавливаются птицами в большем количестве, чем светлые формы, лучше приспособленные к цвету среды. Если же сельскохозяйственные районы станут промышленными центрами, положение изменится коренным образом. В этом случае изменится направление действия естественного отбора, и меланистические формы будут иметь преимущество. В этом случае добычей птиц гораздо чаще будут светлые особи, чем темные. В результате этого ген с подвергается все дальше идущей элиминации из популяции, а ген С широко распространяется.
В предыдущих условиях ген С не мог распространяться в популяции, хотя формы carbonaria являются более устойчивыми. Со времени радикального изменения условий ген С, перед этим элиминируемый, оказался геном, гораздо лучше адаптирующим особей. Поэтому в некоторых промышленных районах меланистические формы очень многих видов ночных бабочек сделались формами, встречаемыми в 90% в отношении к типичным формам и даже в более высоком соотношении.
Можно было бы предполагать, что в конце концов ген с вообще исчезнет из популяции промышленных районов, и лишь отдельные особи будут появляться в виде исключения в результате мутации гена С в ген с. Однако такое положение вероятно никогда не будет иметь места, и в популяции всегда будет встречаться определенный процент светлых форм. Это объясняется тем, что меланистические гетерозиготы Сс являются, как гетерозиготы, более живущими, чем гомозиготы СС, так что в конце установится определенное равновесие частоты проявления обоих аллели (С и с) в каждой популяции.
Мы хотели бы обратить внимание еще на один факт. Он связан с указанной выше переменой в силе доминирования гена С. Спросим себя, почему раньше ген С не был геном полностью доминирующим, как это имеет место в настоящее время? Как известно, свойства организма по существу зависят не от отдельных пар аллели, а от всего набора генов особи. Как каждый ген может воздействовать на большое количество признаков, и наоборот, каждый признак зависит от многих генов, а может быть и от всего набора генов, то есть от целого генотипа. Если вначале ген С не проявлял полного доминирования, то отбор протежировал тем особям, которые благодаря соответствующим генам-модификаторам привели к тому, что у гетерозигот Сс проявлялось более сильное темное окрашивание. В результате селекции, ген, который вначале не проявлял полного доминирования, становился геном полностью доминирующим.
Современная генетика принимает, что гены, вначале рецессивные, если они имеют селекционное значение, превращаются под действием отбора в доминирующие гены. В настоящее время существуют разные гипотезы, пытающиеся выяснить это явление, и возможно, что все они могут быть правильными в зависимости от изучаемого объекта. Однако обычно принимаем, что рецессивный ген, обладающий положительной селекционной ценностью, становится доминирующим геном благодаря соответствующему действию модифицирующих генов, то есть изменяющих действие других генов. Обычно гены, возникшие при мутации, являются рецессивными. Однако это правило не является абсолютным и из него есть исключения.
Обращает на себя внимание тот факт, что все гены, вызывающие меланизм, являются генами частично или полностью доминирующими. Этот факт заставляет предположить, что гены, вызывающие меланизм, появлялись уже не раз, и что они стали доминирующими в результате отбора.
Kettlewell изучал фауну ночных бабочек средней Шотландии, в районах, очень отдаленных от промышленных предприятий. Изучаемые районы покрыты старым лесом, напоминающим леса Великобритании периода нескольких тысяч лет тому назад. Оказалось, что именно в этих условиях удалось обнаружить меланистические формы по крайней мере у семи видов ночных бабочек. Меланистические формы вида Clear a repandata зависят от доминирующего гена и составляют около 10%. Светлых форм этого вида было около 90%.
Светлые бабочки, сидящие днем на коре деревьев, почти невидимы. Однако в этих лесах они в течение дня часто меняют место, потревоженные муравьями или другими насекомыми. В полете светлые бабочки в темном лесу лучше заметны, чем темные, и тогда легче становятся добычей птиц. Это одна из причин, по которой меланизм мог быть раньше полезным. Она может также объяснить равновесие, существующее между светлыми и темными формами в лесах Rannoch. В этом случае меланизм мог сохраниться и затем распространится в результате развития промышленности".
Меланизм встречается также в районах, совершенно лишенных промышленных предприятий, как например, Новая Зеландия, в темных и влажных лесах. Другими словами, меланизм - это очень старое явление, так как всегда имелись среды, в которых темные формы оказывались под охраной естественного отбора, обеспечивающего им выживание. В течение длительного времени естественный отбор мог привести к доминированию первично рецессивных генов, и поэтому встречаемый в настоящее время меланизм зависит от доминирующих генов.
Ясно, что такая теория не исключает первичного отсутствия меланизма у многих видов, которые приобрели его благодаря первичным мутациям. Во всяком случае факт распространения меланистических форм в зависимости от внезапного изменения условий окружающей среды показывает нам зависимость естественного отбора от конкретных условий среды. Этот пример является поучительным и с другой точки зрения. Незначительное изменение окраски может иметь большое значение как адаптационный признак.
Выяснение этого явления стало возможным благодаря объединению генетических и экологических исследований, занимающихся всеми связями, существующими между данной популяцией и совокупностью факторов среды, как живой, так и мертвой. В нашем примере следовало принять во внимание не только роль птиц, но и влияние осадков на лишайники, а также совокупность изменений, вызванных развитием промышленности в данном районе. Кроме того в приведенном примере мы видим, как изменение окраски происходило во взаимосвязи с целым рядом других физиологических изменений, изменений в развитии и поведении, причем по существу не имело значения, шла ли речь о плейотропном действии одного гена, или же о действии различных аллелей, тесно связанных друг с другом.
Приведенный пример относится к так называемому преходящему или временному полиморфизму. Полиморфизмом по Форду называем появление в одном районе в одно и то же время двух или большего количества разных форм одного и того же вида. Частота появления этих форм должна быть большей, чем это можно было бы объяснить происходящими мутациями.
Преходящим полиморфизмом мы называем такое состояние, когда от момента внезапного изменения условий резко увеличивается частота проявления гена, который в прошлом был очень редким или обязан своим происхождением мутации. У некоторых видов ночных бабочек ген темной окраски встречается в небольшом количестве, но с изменением условий, то есть с развитием промышленности в данном районе, приобретает большую селекционную ценность и быстро распространяется. У некоторых видов в старых условиях этот ген вызывал проявление такого вредного признака, что быстро элиминировался под влиянием естественного отбора. Когда же условия изменились, появляющиеся новые мутации стали полезными и в течение короткого времени, нескольких десятилетий, смогли распространиться в популяции, элиминируя когда-то полезные аллели.
Можно предположить что в конце концов прежний ген, вызывающий светлую окраску, будет целиком удален из популяции. Однако может случиться, как мы уже указывали, что этот ген не будет полностью отброшен, если гетерозиготы отличаются определенными положительными признаками. В этом случае со временем установится равновесие между частотой проявления гена светлой и темной окраски в популяции.
Однако независимо от того, каков будет окончательный результат действия отбора, происходящая эволюционная перемена в какой-то популяции заключается в изменении частоты проявления определенных генов. Другими словами, в процессе эволюции происходит изменение генофонда каждой популяции. Это может быть количественное или качественное изменение. Если изменения будут касаться только частоты проявления генов, то мы говорим о количественных изменениях. Если же изменения будут также заключаться в сохранении путем действия отбора мутационных изменений, то есть таких генов, которых раньше не было в популяции, то мы говорим также о качественных изменениях. Ввиду того, однако, что весь набор генов действует как одно целое, на что мы уже указывали выше, действие отбора заключается в селекции наборов генов, а не отдельных генов.
Перейдем к следующему примеру. Объектом изучения в этом случае в течение ряда лет была бабочка (Euphydryas aurinia) относящаяся к русалкам (Nymphalni). Речь идет о популяции, заселяющей изолированный болотистый район. Исследования начались в 1917 г., однако мы располагаем данными уже от 1881 г., о проявлении этого вида в описываемом районе.
В 1881-1897 годах этот вид, производящий в течение года только одно поколение, встречался в большом количестве. Потом, однако, встречался все реже, и с 1917 г. взрослые формы относились к редкостям. Немногочисленная популяция отличалась постоянством признаков и не проявляла почти никакой изменчивости.
Внезапно в 1920 г. наступило резкое количественное увеличение популяции, которая увеличивалась численно до 1924 г. В это время, между 1920 и 1924 годами, популяция отличалась очень большой изменчивостью, касающейся величины бабочек, их окраски и формы цветных пятен. Некоторые особи изменялись до такой степени, что с трудом летали или вообще не были способны летать. С момента, когда количество бабочек стабилизировалось, то есть с 1925 года, бабочки снова стали мало изменчивыми, причем они уже отличались от бабочек до 1920г., то есть от тех, которые жили перед появлением усиленной изменчивости.
Форд объясняет это явление следующим образом. Условия жизни никогда не бывают совершенно одинаковыми. В результате этого и сила, с какой действует естественный отбор, неодинакова. Раз отбор действует сильнее, затем действие его ослабевает. Когда в результате действия условий среды ослабнет действие отбора, в популяции могут появиться такие комбинации генов, которые раньше тщательно отбрасывались. Поэтому не только увеличивается количество популяции, но и одновременно изменчивость ее.
Когда снова, в связи с определенной временной стабилизацией условий, заостряется действие отбора, привилегированными остаются те комбинации генов, которые в настоящее время являются наиболее соответствующими. Количество особей в популяции не только уменьшается, но одновременно особи в популяции становятся мало изменчивыми и выглядят иначе, чем раньше. При стабилизации условий на более длительный промежуток времени, появляются условия для того, чтобы отбор привел к образованию прочной связи тех наборов генов, которые оказываются наиболее соответствующими в данных условиях. Наступает определенное равновесие, которое продолжается до тех пор, пока условия радикальным образом не изменятся.
Многие исследования на лоне природы были проведены над количественными признаками, зависящими от набора полимерных генов. Бабочки, относящиеся к виду Maniola jurtina, отличаются разным количеством темных пятнышек на задних крыльях. Количество пятнышек колеблется от 0 до 5. Оказалось, что в разных местностях живут популяции, отличающиеся по этому признаку.
Казалось бы, что количество пятнышек является совершенно безразличным признаком для выживания особи и ее плодовитости. Оказалось, однако, что полимерные гены, влияющие на количество пятнышек, должны оказывать также плейотропное действие, имеющее разное селективное значение в зависимости от условий, в которых эти формы живут. Поэтому на мелких островах, на которых условия не являются одинаковыми, встречаются популяции, отличающиеся средним количеством этих пятен. Исследования показали, что если условия изменяются, изменяется и направление действия отбора и изменяется формирование полигенических признаков.
Очень интересные эксперименты над действием отбора произвели австралийские ученые под руководством A.J. Nicholson на мухе Lucilia cuprina, личинки которой живут в мясе и ранах овец. Эти эксперименты были произведены в лаборатории.
В качестве питательного материала ученые пользовались растертой говяжьей печенью. В эксперименте изменяли количество пищи, которое получали личинки и зрелые насекомые. Ученые убедились, что уменьшая количество корма личинкам, можно было привести к возникновению штамма мух, обходившегося таким количеством корма, которого было недостаточно для первичных культур.

Рис. 59. Бабочка Maniola jurtina, с брюшной стороны. На другой паре крыльев сбоку видны два темных пятнышка; по W.H. Dowdeswell
Нормальные зрелые мухи, чтобы они могли откладывать яички, должны питаться мясной пищей. Однако путем отбора в определенном направлении выведены породы мух, которым вообще не давали мясной пищи, а только воду и сахар. Кроме того замечено, что если мухи первичных культур откладывали яички только на мясе, то соответственно селекционированные мухи откладывают яички без предыдущего соприкосновения с мясом. В относительно короткий промежуток времени удалось у этого вида мух вывести линию, значительно отличающуюся своими свойствами от первичных пород.
В подробно приведенном примере меланизма ночных бабочек мы имеем дело с преходящим полиморфизмом. Известны также другие формы полиморфизма, с которыми следует познакомиться, так как они наглядно показывают, каким образом может действовать естественный отбор.
В некоторых случаях полиморфизм обусловлен действием факторов окружающей среды. Признаки, как мы уже не раз подчеркивали, зависят как от генотипа, так и от условий среды. Естественный отбор может привести к тому, что та же особь будет обладать разными признаками в зависимости, например, от времени года, если это имеет селективное значение.
Многие млекопитающие животные, живущие на далеком Севере, изменяют окраску своего мехового покрова в зависимости от времени года. Летом они имеют более серый или бурый цвет, тогда как зимой они бывают белого цвета. Одна и другая окраска является покровительственной в зависимости от сезона. В этом случае мы имеем дело не с истинным полиморфизмом, так как обе формы, бурые и белые, никогда не встречаются одновременно.
Если однако обе разные формы появляются одновременно, используя разные среды, то есть экологические ниши, причем различия между этими формами зависят от условии окружающей среды, то есть являются по существу реакцией того же самого генотипа на влияние среды, то мы говорим о полиморфизме, обусловленном средой. Такие примеры дают нам бабочки.
Неподвижные куколки легче всего могут стать добычей врагов. В умеренном климате чаще всего имеются две среды для куколок бабочек, отличающиеся по цвету. Одна среда имеет коричневый фон, другая - зеленый. Куколки тех бабочек, которые переживают эту стадию на земле или стволах деревьев, бывают обычно овальной формы, гладкие, коричневого цвета, Те же, которые проходят эту стадию между листьями деревьев, не имеют такой правильной формы и бывают зеленого цвета. Оказалось, что один и тот же вид бабочек может давать куколки обоих видов. Куколки бабочки махаон (Papilio machacn) находящиеся на стволах, чаще всего бывают коричневого цвета, тогда как куколки на листьях бывают зеленого цвета.
Исследования показали, что кроме самого цвета среды и другие факторы входят в игру, тем не менее "ясно, что под влиянием естественного отбора возникла такая генетическая конституция, которая стремится к созданию зеленых куколок на зеленом фоне и коричневых на коричневом" (P.M. Sheppaid). Такое же явление отмечено и у других видов бабочек.
Следующим видом полиморфизма, зависящим от наследственных факторов, является стойкий полиморфизм, в котором обе формы встречаются в постоянном количественном взаимоотношении, причем это взаимоотношение не изменяется, как мы это наблюдали в преходящем полиморфизме. Теоретически, если в генофонде какой-либо популяции имелись бы две аллели одинакового селекционного значения, то частота их проявления не должна была бы изменяться, как это вытекает из закона Харди-Вайнберга.
Назовем эти аллели А и а. Тогда в популяции имелись бы генотипы АА, Аа и аа, и то в определенном взаимоотношении. Трудно, однако, предположить, чтобы эти все три разные генотипы имели одинаковое селекционное значение. Все наблюдения противоречат такому предположению. Кроме того, если вначале в генофонде популяции имелся только аллель А, который, благодаря мутации, дал аллель а той же селекционной ценности, то распространение этой аллели в популяции происходило бы очень медленно. Вывод этот сделан на основании математического анализа.
Обычно стойкий генетический полиморфизм, с каким мы встречаемся довольно часто, должен быть основан на других принципах. Он возникает в результате определенного равновесия селекционных факторов, благодаря которым ни одна форма не элиминируется полностью. "Это имеет место тогда, когда наиболее редкий из аллелей всегда является полезным, причем по мере того, как он становится все более частым, его селекционное значение уменьшается и в конце концов становится отрицательным" (Sheppard). Такое явление называется сбалансированным полиморфизмом. Чаще всего оно встречается тогда, когда гетерозиготы являются более ценными, чем обе гомозиготы.
Рассмотрим предыдущий пример. В популяции встречаются аллели А и а. Допустим, что, в крайнем случае, как гомозиготы АА, так и аа являются летальными, то есть имеют отрицательную селекционную ценность. Зато гетерозиготы Аа живучи и плодовиты. В результате скрещивания между собой гетерозигот Аа появляются генотипы: 25% АА, 50% Аа и 25% аа.
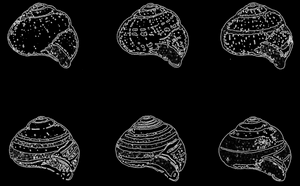
Рис. 60. Улитка Cepaea nernoralis. Объяснение в тексте; по W.H. Dowdeswell
Гомозиготы АА и аа гибнут или не производят потомства, а гетерозиготы Аа размножаются. В каждом поколении оба вида гомозигот элиминируются, а в размножении принимают участие только гетерозиготы. Ясно, что если условия не изменятся, то удержится одинаковое количество аллелей.
Гораздо чаще, однако, дело обстоит иначе. А именно, в том случае, если оба вида гомозигот являются только в некоторой степени менее полноценными, чем гетерозиготы. Если, например, гетерозиготы Аа отличаются лишь несколько большей плодовитостью, чем гомозиготы АА, в популяции неизбежно увеличится количество генов а, хотя гомозиготы аа являются в селекционном значении менее ценными, чем гомозиготы АА. Потеря генов а гомозиготами аа уравновешивается с излишком большей плодовитостью гетерозигот Аа.
Когда количество генов а в популяции увеличивается еще больше, то в конце концов наступает равновесие между потерей генов а гомозиготами и увеличением количества этих генов гетерозиготами. Таким образом, через некоторое время наступает равновесие между количеством аллели А и в популяции, которое сохранится до тех пор, пока не изменятся условия. Описано относительно большое количество таких примеров сбалансированного полиморфизма, который встречается также в популяции людей.
Хорошо разработан случай уравновешенного полиморфизма среди форм улиток (Cepaea nemoralis). Разные формы отличаются как по цвету раковины, так и по расположению на ней темных полосок (рис. 60). В основном встречается раковина трех цветов: желтого, розового и коричневого. Коричневый цвет преобладает над розовым и желтым, а розовый - над желтым. Признак - отсутствие полос доминирует над признаком их наличия. Если у улиток имеются полоски, то их обычно бывает 5. Особи с тремя полосками являются доминирующими в отношении к формам с пятью полосками. Оба признака: цвет скорлупы и полосы зависят от разных, не сцепленных друг с другом пар аллели.
Шеппард и Кейн (Cain) обнаружили, что в лесах с однообразным темным покровом преобладают формы с коричневой или розовой раковиной, без полос или с одной полосой. На зеленом покрове, на траве преобладают формы с желтыми раковинами и пятью полосами. Главным врагом этих улиток является певчий дрозд (Turdus ericetorum). Изучая в разных средах остатки раковин улиток, уничтоженных птицами, можно убедиться, что коричневый и розовый цвет является защитным для улиток в лесах с коричневым покровом, а желтый - на травостоях и среди живых изгородей.
Почему, однако, несмотря на селекционное значение покровительственной окраски, на коричневом покрове встречается определенный процент улиток с желтыми раковинами а на зеленом покрове - с розовыми и коричневыми? Взаимоотношения являются гораздо более сложными, чем могло бы казаться. Вероятно имеет значение большая плодовитость гетерозигот, чем можно объяснить сложные полиморфические отношения в популяциях Cepaea nemoralis, обитающих в разных средах.
Некоторые формы полиморфизма, встречающиеся у человека, и касающиеся проявления разных аллелей, предопределяющих группы крови, а также аллелей, обуславливающих способность человека различать горький вкус химического соединения - фенилтиомочевины, возникли в очень давние времена. Аналогичные примеры полиморфизма встречаются у современных человекообразных обезьян, что указывает на возникновение этого вида полиморфизма еще у общих предков человекообразных обезьян и человека, то есть несколько десятков миллионов лет тому назад, в миоценовую эпоху. В других случаях сбалансированный полиморфизм существует по всей вероятности уже тысячи лет.
Как обнаружил Diver, разные формы полосатости раковины Cepaea встречаются в плейстоцене. Так как виды, находящиеся в близком родстве, имеют почти одинаковые полиморфные разновидности, можно предполагать, что их общий предок уже обладал таким полиморфизмом. Иногда само явление полиморфизма оказывается более стойким, чем вид или даже более высокая систематическая единица.
Кончая данную главу, мы хотим еще раз подчеркнуть те пункты, которые, как нам кажется, являются самыми главными.
Оказывается, что единицей, на которую действует естественный отбор, является популяция. Особь сама по себе исполняет второстепенную роль, хотя популяция состоит из особей. В популяции Biston отдельные меланистические особи в промышленных районах могут сделаться добычей птиц, а могут выжить и оставить потомство. Однако в сумме темные особи оставляют больше потомства, чем хуже приспособленные светлые. В результате этого генофонд популяции постепенно изменится. Общий генофонд будет обогащаться аллелями, вызывающими темное окрашивание, и терять аллели, вызывающие светлое окрашивание.
Таким образом, в результате отбора изменяется фонд генов популяции, изменяется частота отдельных генов, могут распространиться гены, которые до того времени удалялись в процессе отбора. Условием этих изменений является изменение условий окружающей среды. Если бы не развилась промышленность, меланистические формы относились бы к редким исключениям, то есть к аберрациям, как их называет систематика.
Развитие генетики с одной стороны и знакомство с цитологическими процессами мейоза и оплодотворения с другой, позволили более точно определить, чем по существу является естественный отбор. Дарвин принимал изменчивость и наследование изменчивости как несомненный факт. В настоящее время мы однако знаем, какой вид изменчивости является наследственным и каковы источники наследственной изменчивости. Кроме того, установление корпускулярной сущности наследственности ликвидировало те трудности, с которыми встретился Дарвин, принимая слитную наследственность.
Учитывая современный уровень знаний, мы можем определить естественный отбор как процесс, всегда имеющий место при половом размножении и являющийся его обязательным атрибутом. Отбор не является какой-то силой. Это совокупность процессов, происходящих внутри популяции, которая подвергается изменениям (Goudge). Благодаря процессу мутации в каждой популяции накапливаются разные аллели, появляющиеся в результате мейоза и оплодотворения в самых разнообразных комбинациях. Разные комбинации генов имеют разную селективную ценность, то есть подвергаются действию отбора. Направление отбора определяется господствующим в данном периоде комплексом условий как неживой, так и живой природы.
Естественный отбор ничего не может предвидеть, действует лишь относительно настоящего момента. Поэтому прав Добржанский, когда говорит об оппортунизме естественного отбора. В этом также заключается основное различие между действием естественного и искусственного отбора, которым управляет человек, согласно принятому плану.
Человек отбирает для дальнейшего разведения формы, наиболее соответствующие его намерениям. Другим особям он не дает возможности размножаться. Природа поступает иначе. Она безжалостно элиминирует только те особи, которые или вообще гибнут, не достигнув половой зрелости, или являются бесплодными. Другие же размножаются, но окончательными победителями оказываются те, которые размножаясь более интенсивно, тем самым способствуют изменениям общего генофонда.