
Вид и видообразование
Хотя главный труд Дарвина называется: "О происхождении видов" многие авторы подчеркивают, что это название не соответствовало полностью содержанию. Труд в основном занимается эволюционными изменениями и механизмом этих изменений, а не видом, как таковым, и его происхождением. Поэтому более соответствовало бы название, например, "Принципы эволюции". С другой стороны, лишь при помощи генетики стал возможным более близкий анализ понятия вида. Как известно, однако, вторичное открытие законов Менделя и развитие генетики наступило спустя много лет после смерти творца теории естественного отбора.
Вид не является иерархически наиболее низкой системной единицей. В виде различаем разновидности, породы и подвиды. Согласно Дарвину, единственный показатель при определении, является ли данная форма разновидностью или видом - это мнение опытных натуралистов, обладающих определенными знаниями. Ничего удивительного, что в "Происхождении видов" встречаемся со следующим высказыванием: "Я считаю, что понятие "вид" употребляется произвольно к совокупности особей, сходных между собой, и понятие это в основном не отличается от понятия разновидности, которым мы определяем формы менее различные и менее изменчивые''.
Как мы видим, Дарвин не отмечал принципиальной разницы между видом и разновидностью. Он считал, что разновидности являются эволюционно новыми видами.
Тем временем человек издавна замечал в окружающей его природе формы, отчетливо отличающиеся от других, которые назвал иначе. Несмотря на общее сходство, лев отличается от тигра, ягуара, рыси и всех других кошек. Между этими различными формами животных, относящихся к семейству кошачьих, в природе никогда не замечено переходных форм, которые являлись бы результатом их взаимного скрещивания.
Иначе дело обстоит с подвидами или породами. Несмотря на значительное иногда различие, эти формы могут скрещиваться между собой в природе, давая в результате промежуточные формы. Можно принять, что между подвидами, относящимися к одному и тому же виду, в природе не существует какой-то преграды, противодействующей их скрещиванию, тогда как такие преграды существуют между особями, относящимися
Две расы, занимающие отдельные территории, мы называем аллопатическими расами. С момента, когда эти две расы превратятся в отдельные виды, они оказавшись вновь на общей территории, сохранят свои особенности благодаря полной генеративной изоляции. Такие виды, заселяющие одну и ту же территорию, носят название симпатрических видов.
Многие авторы принимают, что расщепление одного первичного вида на два или больше видов может наступить только в результате преобразования географических рас, то есть генетически открытых систем, в генетические закрытые, благодаря географической изоляции. Некоторые из биологов однако считают, что даже популяция, заселяющая одну и ту же территорию, может расщепиться на две отдельные популяции, специализирующиеся в освоении другой пищи, размножении в другое время и так далее. Эти авторы считают, что на той же территории могут возникнуть две расы, которые со временем могут дать начало двум симпатрическим видам.
Дарвин, как известно, обращал внимание на значение дивергенции для процесса эволюции. По его мнению, формы, наиболее отличающиеся друг от друга, имеют больше всего шансов выйти победителями в борьбе за существование, так как могут использовать окружающую среду другим путем, и таким образом избежать соперничества с другими формами. Однако нам кажется, что хотя бы вначале, взаимное скрещивание особей одной популяции привело бы к нивелировке генетических различий.
Nicholson считает, что в начальной фазе необходим изоляционный барьер, который сделает возможным дифференциацию монолитной первоначально популяции на две, под влиянием действия отбора. Если в результате этой изоляции обе популяции приспособятся к разному использованию окружающей среды, и при этом будут настолько изолированы друг от друга генеративно, что помеси не будут обладать такой же плодовитостью, как потомки двух особей, относящихся к той же расе, естественный отбор может привести к полной генеративной изоляции.
Действие отбора в этом направлении доказано экспериментальным путем. Koopman производил исследования на двух видах дрозофил, находящихся в близком родстве - Drosophila pseudoobscura и Drosophila persimilis. В естественных условиях в природе почти никогда не происходит скрещивания между особями, относящимися к этим двум видам. При искусственном разведении, особенно при пониженной температуре, могут легко происходить межвидовые скрещивания. Гибриды являются совершенно бесплодными. В условиях разведения помещали вместе одинаковое количество самцов и самок обоих видов. Гибриды, которые являются бесплодными, как мы уже указывали выше, отбрасывали при дальнейшем разведении. Вначаее количество этих гибридов было большим, но затем это количество постепенно уменьшилось. Все чаще происходила копуляция мух, относящихся к одному и тому же виду.
Результаты этого опыта можно объяснить следующим образом. Вначале в популяции обоих видов дрозофил появляются особи, отличающиеся друг от друга генетически с точки зрения сексуальной разборчивости. Ясно, что мы имеем ввиду только условия разведения, так как в природе не происходит межвидового скрещивания. В условиях искусственного разведения, однако, одни мухи охотно скрещивались с мухами другого вида, тогда, как другие подбирают себе партнеров того же самого вида. Ввиду того, что гибриды являются бесплодными, мухи, скрещивающиеся с представителями другого вида, не передают своих генов потомству. Мухи же, которые скрещивались с особями того же вида, могли передать свои наклонности, обусловленные генетически, следующим поколениям. Наступил процесс селекции, приводящий ко всё дальше идущей элиминации особей, скрещивающихся с особями другого вида, то есть, другими словами, селекция приводит ко все более четкой генеративной изоляции обоих видов.
Аналогичный эксперимент произвели другие авторы. Они разводили вместе одинаковое количество дрозофил, относящихся к двум разным рецессивным мутациям. Обе расы были гомозиготами и отличались фенотипами, которые легко отличались друг от друга.
Обозначим рецессивный ген одной расы а, рецессивный ген другой расы - b. Нормальными доминирующими аллелями этих генов у диких мух являются гены А и В. Мухи одной расы имеют генотип ааВВ, мухи другой расы - AAbb. В результате скрещивания этих рас возникают гетерозиготы АаВb, обладающие генотипом диких мух, которые легко отличить от мух ааВВ и AAbb. Из эксперимента отбрасывали всех мух с фенотипом диких, то есть гетерозигот АаВb. Оказалось, что с течением времени число возникающих гетерозигот уменьшается, то есть что начинает образовываться механизм, изолирующий генеративно обе исходные расы мух.
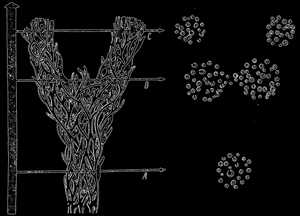
Рис. 64. Схема видообразования. Объяснение в тексте; по Т. Добржанскому
Процесс видообразования лучше всего отображает схема, взятая из книжки Добржанского (рис. 64). Переплетенные ветки как бы образуют общий ствол. Каждая веточка представляет одну популяцию. Отдельные ответвления, соединяющие ветви, обозначают обмен генов между популяциями, которые не изолированы генеративно друг от друга. В течение длительного времени на приведенной схеме до уровня, обозначенного линией А, все популяции этого вида генеративно мало дифференцированы. Зато в промежутке времени, заключенном между линиями А и В, можно заметить, что совокупность популяций начинает разделяться на две группы, представляющие две возникающие географические расы. Поперечный разрез через популяцию вида дает на уровне А картину сгруппированных друг с другом популяций, что свидетельствует о незначительном различии между популяциями. Разрез на уровне В даст уже другую картину.
Мы видим, что вид распался на две расы, каждая из которых состоит из большого количества популяций. Количество промежуточных популяций между двумя расами довольно невелико. Глядя на уровень В можно предвидеть, что случиться в следующем периоде, заключенном между линиями В и С. Монолитный вначале вид разделиться на два отдельных вида, изолированных генеративно друг от друга. На разрезе увидим две отделенные друг от друга популяции, относящиеся, уже к двум видам.
Приведенная схема помогает нам понять процесс видообразования. На уровне В процесс разделения двух рас зашел так далеко, что обе расы постепенно набирают признаки отдельных видов.
Для нас особенно наглядными являются те примеры в природе, которые как бы in statu nascendi иллюстрируют нам процесс видообразования, некоторые из этих примеров следует разобрать. Один из них относится к лягушке Rana pipiens из Соединенных Штатов. Этот вид распространен от северных штатов до Флориды и Техаса. Географические формы этого вида, живущие по соседству, легко скрещиваются друг с другом и дают плодовитое потомство. Если же в эксперименте скрестить северных лягушек с южными, наступает оплодотворение, зигота вначале развивается, но затем зародыши гибнут. Оказалось, что северные и южные формы изолированы генеративно.
В природе они по существу никогда не имеют возможности скрещивания, однако ввиду того, что на всем пространстве от севера до юга встречается тот же вид лягушек, обмен генов между отдельными формами может происходить очень постепенно. Однако обмен генов происходит так медленно, что он не затормозил постепенно развивающейся генеративной изоляции.
Предположим, что по какой-то причине наступает географическая изоляция северной расы от южной. В этом случае мы не сомневались бы в том, что северные и южные формы относятся к разным видам. В настоящее время, так как обе расы соединены рядом переходных форм, мы, правда, относим северные и южные формы к одному виду, однако этот вид все отчетливее дифференцируется f на два отдельных вида. На приведенной выше схеме вид Rana pipiens следовало бы расположить на линии В или несколько выше ее.
Между северными и южными формами имеется некоторое различие. Яички южных форм, например с Флориды, гораздо мельче, хуже переносят более низкие температуры среды и лучше - более высокие.
Другой пример относится к двум видам чаек. На Британских островах и в Скандинавии жидут рядом друг с другом, два вида: Larus argentatus и Larus fuscus. Хоть они и живут на той же территории, но никогда не скрещиваются друг с другом и между ними нет переходных форм. Как обнаружил Майр (Мауг), оба вида объединяются целым рядом географических рас, образующих непрерывную цепь вокруг Ледовитого океана, Лабрадора, Канады, Северовосточной и Северозападной Сибири и северной России.
На рисунке 65 представлен этот ряд разновидностей, заселяющих разные территории. На территории, занятой двумя крайними расами, различия между ними уже носят характер генеративной изоляции. Принимая во внимание очередные расы, скрещивающиеся между собой, мы можем обе формы L. argentatus и L. fuscus назвать географическими расами, а принимая во внимание их генеративную изоляцию - отдельными видами.
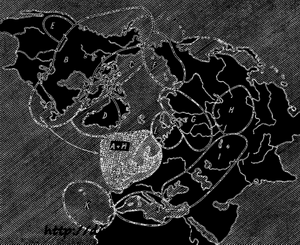
Рис. 65. Географическое размещение чаек в роде Larus. Две формы, не скрещивающиеся друг с другом, населяют в настоящее время одну территорию (А + М). Эти формы соединены рядом рас, живущих по соседству, (В, С, D, F, G, и L), которые скрещиваются между собой по Т. Добржанскому(А + М). Эти формы соединены рядом рас, живущих по соседству, (В, С, D, F, G, и L), которые скрещиваются между собой по Т. Добржанскому">
На территории Британских островов и Скандинавии невозможен обмен генов между L. argentatus и L. fuscus. Оба эти вида могут обмениваться генами косвенным путем, через отдельные звенья цепи географических рас. Однако этот обмен происходит так медленно, что он не мог противодействовать образованию генеративной изоляции между крайними формами.
Следующий интересный пример относится к хвостатым земноводным, живущим в Калифорнии. Они живут в горах, окружающих непрерывным кольцом горячую и сухую низменную пустыню, лежащую среди этой горной цепи. В низменности этих животных нет совершенно.
Как доказал Стеббинс (R.С. Stebbins), вид Ensatina eschscholtzi встречается в зависимости от зоны в разных географических расах. Отдельные расы отличаются друг от друга окраской, а живущие по соседству расы скрещиваются друг с другом и между ними происходит обмен генов.
Типичная форма eschscholtzi из западной и южной Калифорнии переходит на севере в форму xanthoptica в районе залива Сан-Франциско, и в форму picta и oregonensis на севере от Сан-Франциско, а затем platensis в горах Сьерра-
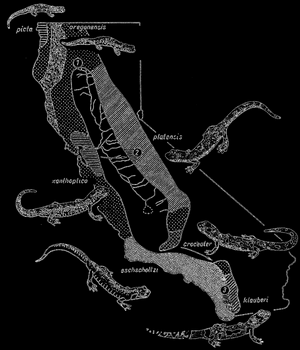
Рис. 66. Видообразование в роде Ensatina. Объяснение в тексте; по Т. Добржанскому
Невада и расы croceater и klauberi (рис. 66). Это все географические расы. Но в южной Калифорнии три наиболее отличающиеся друг от друга формы, а именно eschscholtzi, eroceater и klauberi живут в одном районе, не скрещиваясь между собой. Их можно было бы считать отдельными видами, однако эти виды объединяются рядом переходных рас, скрещивающихся друг с другом.
Хотя центральная низменность является изоляционным барьером между расами, живущими на запад и восток от нее, форма xanthoptica преодолела эту низменность, и ее можно найти также в районе, заселенном расой platensis. Эти расы могут скрещиваться между собой, то есть в этом районе еще не существует генеративной изоляции между этими расами, тогда как на юге такая изоляция между живущими там расами уже существует.
Гребенчатый тритон (Friturus cristatus) встречается в многочисленных географических подвидах. Скрещивая между собой формы, живущие в окрестностях Неаполя и Лондона, получаем около 80% нормально развивающихся личинок. Во втором поколении, однако, лишь 4% личинок способны к нормальному развитию. Это является результатом расстройства в конъюгации хромосом гибридов.
Приведенные примеры указывают на трудности в отнесении некоторых форм к отдельным расам или отдельным видам. Примеры этого характера представляли большие трудности для давних систематиков, принимающих как правило постоянство и неизменяемость видов. С принятием эволюции и динамического, а не статического понятия вида, эти факты являются одним из важных доказательств эволюционного процесса в природе.
Видообразование в настоящее время является фактом, причем выяснение его процессов обосновано на тех же принципах, на которых обосновано явление микроэволюции. Так, по крайней мере, считает огромное большинство исследователей. Другими словами, как микроэволюцию можно объяснить действием естественного отбора, так и видообразование обусловлено действием того же самого фактора.
В течение последних десятилетий систематики все больше внимания посвящают вопросу так называемых - видов близнецов. Это два, три или больше видов, почти идентичных морфологически, которые в естественных условиях проявляют полную генеративную изоляцию. В тропической Америке, например, встречается пять видов дрозофил, которые лишь в лабораторных условиях могут в исключительных случаях скрещиваться друг с другом, давая бесплодное потомство. Эти виды так сходны между собой, что лишь недавно обнаружено между ними минимальное различие в наружных копулятивных органах самцов и различие в хромосомах.
Аналогичные примеры близнецовых видов встречаем и у других организмов, в том числе и у инфузорий. Так, например, среди представителей старого вида Paramecium aurelia, Соннеборн (Sonneborn) и другие обнаружили несколько подвидов, генеративно изолированных друг от друга. Особи, относящиеся к разным подвидам, или вообще не конъюгируют друг с другом, или же в результате их конъюгации возникают особи, неспособные к жизни и размножению.
Правда, между некоторыми разновидностями отмечаются незначительные различия, например, по величине, однако, как правило, лишь анализ половых процессов, то есть конъюгации, и поведения особей после конъюгации, может быть единственным верным показателем того, следует ли отнести исследуемые формы к той или другой разновидности.
Как видим, отдельные разновидности Paramecium aurelia являются хорошими видами с точки зрения систематики. Для облегчения классификации им однако не дают специальных видовых названий, а обозначают как разновидности 1, 2, 3 и так далее. Близнецовые виды несомненно произошли эволюционным путем из общего ствола. Процесс видообразования может у разных форм происходить по-разному. Пока неизвестно, как он происходит, например, у туфелек.
Если при помощи биологических и генетических критериев у форм, размножающихся половым путем, можно ближе уточнить вид, как генетически замкнутую систему, то эти критерия оказываются бессильными, если мы имеем дело с видами, размножающимися исключительно вегетативным, то есть бесполым путем (путем деления, почкования и так далее). Правда, число видов, которые никогда не размножаются половым путем, или у которых не происходит рекомбинации генов, невелико, но нельзя отрицать, что формы, размножающиеся исключительно вегетативно, существуют. Так, например, бактерия, размножающаяся делением, даст в результате ряды особей такого же самого генотипа, то есть так называемые клоны. Изменение генотипа может произойти лишь в результате мутации.
У форм, размножающихся исключительно бесполым путем, трудно по существу говорить о виде. Назовем ли мы изменения, возникшие среди них в результате мутации, разновидностями, видами, или даже родами - это условное понятие. Поэтому разные авторы по-разному классифицируют эти формы, и в этом случае невозможно ожидать быстрого и полного согласования взглядов. Это также относится к разграничению видов растения из рода Crataegus и Ribes.
В этом случае мы имеем дело с так называемой апомиксией, при которой половое размножение заменено таким видом размножения, где не имеет места ни мейоз, ни оплодотворение. У животных иногда встречается размножение в виде партеногенеза.
Как мы уже не раз подчеркивали, естественный отбор лишь тогда имеет место, когда имеет в своем распоряжении определенный запас наследственной изменчивости. Первичным источником наследственной изменчивости являются разного рода мутации, о которых учит генетика. Однако ввиду того, что в каждой популяции имеются разные разновидности, то есть аллели почти каждого из генов, возникших при мутации, в результате чего возникают гетерозиготные особи в многих парах аллелей, то благодаря мейозу и опло-дотверению в каждый момент существует соответствующий запас наследственной изменчивости, который может служить материалом для естественного отбора. Таким образом, у существ, размножающихся половым путем, отбор может действовать, хотя в течение длительного времени не появлялось никаких новых мутаций.
Генофонд каждой популяции очень разнообразен, благодаря обильному запасу разных аллелей. Поэтому если монолитная вначале популяция будет разделена каким-то барьером на две части, из которых одна состоит лишь из небольшого числа особей, то эти особи не могут содержать в своих генотипах всех аллели, встречающиеся в первичной многочисленной популяции. Это является делом чистой случайности, которые аллели окажутся в группе особей, отделенных географическим барьером.
Предположим, что в результате вулканической деятельности из волн океана появится океанический остров. На этот остров могут попасть загнанные бурями с ближайшего континента отдельные представители определенного вида птиц. Это дело случая, какие гены из всего генофонда данного вида птиц окажутся на новообразованном острове. Некоторые гены могут вообще отсутствовать, частота же проявления остальных будет совершенно другой, чем в первичной популяции. На этот генетический субстрат будет и дальше действовать естественный отбор, уже в новых условиях, существующих на острове.
Вообще можно сказать, что дальнейшая эволюция этого вида в большой степени будет зависеть от случайного набора генов, содержащихся в генотипах особей, которые оказались на новой территории, где условия существования отличаются от существовавших на их старой родине. Другими могут быть не только климатические, но и биотические условия.
Особи вида, прибывшие на остров с ближайшего континента, не найдут там своих старых соперников, благодаря чему они могут заселить новые экологические ниши, которые раньше были им недоступны. Этих несколько особей - основоположников, которые случайно поселились на океаническом острове, можно считать единственными основоположниками породы, из которой при дальнейшем развитии сформируются новые виды, называемые эндемическими. Из эндемических видов в свою очередь образуются новые семейства. Это зависит только от времени. Чем дольше длится изоляция, тем более высокие систематические категории могут появиться. Другими словами, в небольших изолированных популяциях судьба эволюционного развития зависит от чистой случайности, и тогда мы говорим о генетико-автоматических процессах значение которого для эволюционных процессов особенно подчеркивал американский генетик С. Райт (S. Wright).
Мнения относительно объема действия и значения генетико-автоматических процессов для эволюционных преобразований пока расходятся. Тогда, как одни авторы считают, что роль этого фактора для эволюции малочисленных популяций очень велика, другие не придают ему никакого значения и считают, что даже в малочисленных популяциях главным фактором эволюции является исключительно естественный отбор.
Правда вероятно находится по середине. Ясно, что при изоляции небольшой популяции от первичной, действие отбора тем самым суживается до генов, представленных генотипами изолированных особей. Поэтому случайный генетический состав изолированной популяции, то есть генетико-автоматический фактор, влияет на дальнейшую ее эволюцию.
Приведем еще один пример. Допустим, что давным-давно с разбитого корабля на необитаемый остров попадает группа людей. Среди них нет представителей всех групп крови. Из групп 0, А, В и АВ имеются только особи группы 0 и А. Особи, относящиеся к группе 0, должны быть гомозиготами, так как ген, вызывающий группу 0, является рецессивным по отношению к аллелям, вызывающим образование группы А и В. Особи, относящиеся к группе А, могут быть гомозиготами АА или гетерозиготами А0. Предположим, что в результате чисто случайных причин новорожденные, содержащие ген А, гибнут. В живых остаются только особи, обладающие геном, вызывающим образование группы 0. Через некоторое время вся популяция этого острова будет состоять из особей, имеющих группу крови 0. Этот результат не вызван действием естественного отбора, а зависит от случая, то есть от генетико-автоматических процессов. (Принятый в русской литературе термин "генетико-автоматические процессы" (Н.О. Дубинин и Д.Д. Рамашов, 1932) соответствует термину "дрейф генов" (Райт, 1931), принятому в западней литературе. (прим. редактора).)
Вообще говоря, не существует противопоставления действия естественного отбора и генетико-автоматических процессов. Последние только ограничивают действие отбора, который должен пользоваться генами, имеющимися в его распоряжении.
Процесс образования новых видов из существовавших раньше является длительным и постепенным, происходящим благодаря накоплению мелких генетических изменений. Это постоянный процесс, приводящий путем образования генетических изменений между популяциями, к возникновению разных рас или разновидностей, а некоторые из них могут в дальнейшем стать отдельными видами.
Некоторые из авторов однако считают, что процесс видообразования может происходить внезапно, скачками. Эти авторы предполагают, что время от времени появляются мутации широкого действия, одномоментно значительно изменяющие всю организацию особи, существовавшую до того времени.
Гольдшмидт называет такие мутации гомеотическими мутациями. В результате внезапно появляются формы, которые no-существу являются как бы уродствами в сравнении с материнскими особями. Ясно что чаще всего эти формы не приспособлены к жизни и гибнут, не оставляя потомства. Однако может случится, по мнению Гольдшмидта, что в результате таких мутаций широкого действия, возникают "уроды с будущим" (hopeful monsters), которые дают начало новой разновидности организмов.
Однако такая теория является совершенно неправильной в свете успехов современной генетики. Во-первых, по существу каждое значительное изменение организма, возникшее в результате мутации, является серьезным минусом. Каждый организм так приспособлен к жизни в господствующих в настоящее время условиях, что значительное изменение является для него в большей или меньшей степени помехой в борьбе за существование. Во-вторых, чтобы урод в будущем мог передать свои признаки следующим поколениям, он должен бы найти соответствующего партнера.
Трудно предположить, однако, чтобы особь, резко отличающаяся от остальных особей, скрещиваясь с особью, резко отличной от себя, могла передать потомству свои признаки. Совершенно невероятным нам кажется, чтобы в то самое время и в том же месте могло мутационным образом возникнуть несколько особей, измененных одинаковым образом.
Поэтому теории, старающиеся объяснить серьезные эволюционные изменения мутациями широкого действия, в настоящее время почти не имеют сторонников. Подавляющее большинство генетиков-эволюционистов принимает, так как принимал Дарвин, что всякие эволюционные изменения происходили постепенно путем накапливания мелких различий в длинном ряду поколений.
Только в исключительных случаях и только у некоторых организмов процесс видообразования может происходить в быстром темпе. Примеры видообразования этой формы встречаются главным образом у растений. Причиной их может быть полиплоидия. Как известно из генетики, различается два вида полиплоидии. Если происходит умножение числа хромосом одного вида (одного и того же исходного набора), то говорим об аутополиплоидах. Этот вид полиплоидии является наиболее частым, и у одного вида растений можно часто встретить формы с увеличением числа хромосом в различное число раз. В этой форме полиплоидии качество генов не изменяется, но аллели, вместе того, чтобы находиться парами, имеются в большем количестве. Изменяется также состояние равновесия между генами и цитоплазмой, что вместе приводит к изменениям фенотипа полиплоидных форм.
Другой формой полиплоидии является аллополиплоидия, при которой умножается число хромосом двух разных видов. Если наступит скрещивание двух разных видов растений, то чаще всего второе поколение гибридов бывает совершенно бесплодным, или часть гибридов бывает бесплодной, а часть обладает большой изменчивостью.
Бесплодие вызвано расстройствами мейоза. Хромосомы не находят партнеров, с которыми могли бы конъюгировать, и в результате появляются гаметы с разными наборами хромосом. Иногда же случается, что хромосомы каждого вида удваиваются в гибриде, и тогда каждая хромосома имеет своего партнера при конъюгации, мейоз наступает правильно и гибрид даже в дальнейших поколениях сохраняет свой характер. Это аллополиплоидия. Таким образом, могут, путем скрещивания двух видов растений, возникать новые виды.
Приведем примеры. Пикульник шероховатый (Galeopsis tetrahit) из семейства губоцветных (Lobiatae) имеет 32 хромосомы, тогда как находящиеся в близком родстве виды Пикульник заметный (G. speciosa) и Пикульник пушистый (G. pubescens) имеют только по 16 хромосом. Шведский генетик Мюнтциг (Muntzig) скрестил оба вида, то есть speciosa и pubescens, и среди гибридов нашел формы с удвоенным числом хромосом, соотвествующие известному из природы G. tetrahit.
Иногда межвидовые гибриды у растений могут появляться без умножения числа хромосом. В этом случае происходит переход генов с одного вида на другой, и тогда мы говорим об интрогрессии. В настоящее время ботаники знают уже довольно многочисленные примеры такой интрогрессии.
Наглядным является пример двух видов ирисов: Iris fulva и Iris hexagona. Оба вида значительно отличаются друг от друга. Iris fulva растет на более сухих грунтах, в лесах, Iris hexagona на влажной почве, не затененной. Человек, вырубая леса и создавая новые почвенные условия, привел к тому, что плодовитые гибриды между двумя первичными видами нашли удобные для себя условия развития.
Аналогичные примеры находим также среди некоторых животных. Плодовитые помеси встречаем, например, между двумя видами жаб, Bufo атеricanus и Bufo fowleri, а также между видами птиц, Pipilo erythrophtalmum и ocai.
Добржанский отмечает, что интрогрессия без умножения числа хромосом возможна, однако, лишь тогда, когда еще не наступило совершенно отчетливое видообразование. Тогда лишь можно получить плодовитые помеси без полиплоидии.
Мюнтцигу удалось, скрещивая 2 вида, создать искусственно новый вид, встречаемый в природе. Он доказывал, что путем межвидового скрещивания иногда могли возникнуть новые виды. Оказывается, что иногда человек таким же путем может создать искусственные виды, которых в природе не встречается.
Советский генетик, Карпеченко, получил путем скрещивания капусты (Brassica aleracea) с редькой (Raphanus sativus) аллотетраплоидный гибрид, который размножался как новый вид. В этом случае растения, взятые для скрещивания, относятся к двум родам - Brassica и Raphanus. Таким образом, была получена новая форма, Raphanobrassica. Как Brassica так и Raphanus имеют по 18 хромосом, Raphanobrassica имеет их 36, то есть является аллотетраплоидом.
Другим путем был искусственно получен новый вид дрозофил, который был назван Drosophila artificialis. Кожевников (1936) вывел из Drosophila melanogaster разновидность, скрещивая две формы этого вида, у которых отмечалось различие хромосом. В этом случае дело заключалось в комбинации двух транслокаций, то есть перемещения во второй и третьей паре хромосом.
Drosophila artificialis образуют четыре формы гамет. Из возможных 16 форм зигот только четыре являются жизнеспособными, другие гибнут в результате различных расстройств (нехваток и удвоений - дупликаций). Эти же четыре сформированы так же, как и родители. Таким образом, Drosophila artificialis с Drosophila melanogaster дает в результате нежизнеспособные гибриды.
Таким образом, была получена искусственная разновидность, бесплодная с материнскими формами.
Описанные выше примеры спорадического возникновения новых видов путем скрещивания относятся, однако, к редкостям, и этот механизм возникновения новых видов несомненно имел и имеет в процессе эволюции очень ограниченное значение. В настоящее время большинство авторов признает, что видообразование является, как правило, процессом, происходящим постепенно, путем накапливания незначительных генетических изменений в пределах популяции.
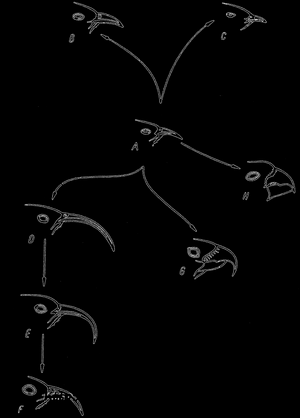
Рис. 67. Эволюция у Drepanidae с Гавайских островов. Вероятно, форма А была исходной формой, из которой развились все другие формы, характеризующиеся прежде всего изменениями формы клюва; по Е.О. DodsonА была исходной формой, из которой развились все другие формы, характеризующиеся прежде всего изменениями формы клюва; по Е.О. Dodson">
По мнению большинства биологов те же факторы, которые приводят к генетическим изменениям популяции, действуют в процессе видообразования, а также и образования высших систематических единиц. Роль играет прежде всего время. Если один вид с течением времени может благодаря искусственному отбору образовать разные генетические популяции и расы, то со временем эти расы могут преобразоваться в разные виды. Другими словами, генетические открытые системы могут превратиться в генетически закрытые. Если, однако, этот процесс эволюционной дифференциации будет иметь в своем распоряжении соответственно длительное время, виды изменятся в отдельные роды, а роды могут образовать отдельные семейства и еще более высокие систематические единицы.
Наверняка не больше, чем несколько особей родом из Америки, дало начало одному семейству птиц (Drepaniidae) с Гавайских Островов (рис. 67). Это эндемическое семейство, известное только на Гавайях, состоит из 18 родов, к которым относится около 40 видов.
Естественное течение эволюционных процессов отображает каждая систематическая классификация. Родственные виды, то есть обладающие сходством, объединяем в один род. Родственные роды объединяем в одно семейство, семейства в отряды, а отряды в классы. Таким образом, каждая классификационная система является именно такой, какой бы следовало ожидать, приняв принцип постепенной эволюции.
В настоящее время большинство биологов принимает, что как в микроэволюции, так и в макроэволюции действуют те же самые факторы и так же, как сегодня, так и в прошедшие геологические эпохи. Таким образом, можно говорить о биологическом униформизме, который со времен Лайеля принят как основа в геологии.
Однако встает вопрос, может ли современная эволюционная теория естественного отбора, базирующаяся на генетических данных, полностью и без остатка объяснить нам эволюционное развитие форм жизни? Нам лично кажется, что в изучении факторов эволюции новые перспективы открывают нам исследования над неправильностью мутаций и преобразованием фено-копий в аналогичные наследственные изменения. Появление определенных мутаций при воздействии определенных условий окружающей среды является фактом, установленным у некоторых микроорганизмов, а прежде всего у дрожжей.
Встает вопрос, не откроет ли наука в будущем стимулов, которые аналогичным образом приводили к образованию определенных мутаций и у других организмов. Нам кажется, что отрицательный ответ в настоящее время был бы преждевременным и не лишенным догматизма. Кроме того, результаты исследований Уоддингтона (Waddington) и его сотрудников указывают на реальную возможность генетического закрепления у поколений вначале не наследственных изменений, то есть фенокопий, вызванных действием на определенном этапе развития определенных стимулов.
Результаты, полученные Уоддингтоном вызвали живую полемику, однако эта проблема нуждается в дальнейших основательных исследованиях, чтобы убедиться, какое значение может иметь факт, открытый и изученный Уоддингтоном. Следует добавить, что впервые на этот факт обратил внимание много лет тому назад немецкий биолог Йоллос (Jollos) затем Гольдшмидт, и в Польше Т. Мархлевски. Для нас наиболее важным является тот факт, что некоторые положения, которые постулировал ламаркизм, могут быть в основном правильными, хотя их современная интерпретация согласована с принципами генетики и селекции, и не основана на принятии принципа наследования приобретенных признаков.
Микроэволюция, приводящая к различиям в генофонде популяции, сделалась предметом тщательного теоретического и экспериментального изучения, а также исследований, производимых в естественных условиях. Действие естественного отбора, о котором писал Дарвин, подтвердилось со всей точностью. Одновременно большинство авторов подчеркивает, что и процессы макроэволюции, охватывающие процессы видообразования, как и дифференциация высших систематических категорий, не нуждаются в действии каких-то дополнительных факторов, кроме тех, которые управляют процессами микроэволюции. Именно такой вывод делает современная теория естественного отбора, или, как ее некоторые называют, синтетическая теория эволюции.