
Родословная человека
В предыдущей главе мы указывали, согласно принятым в настоящее время взглядам, что в третичном периоде, вероятней всего в миоценовую эпоху, какое-то первобытное племя человекообразных обезьян разделилось на две линии. Одна из них привела в эволюционном развитии к человеческим формам, вторая - к живущим в настоящее время человекообразным обезьянам. Встает вопрос, который из континентов являлся колыбелью человеческого рода, на каком континенте наступили те эволюционные изменения, которые имели и имеют столь огромное значение для дальнейшей судьбы жизни на нашем земном шаре?
Дарвин считал, что этим континентом была Африка. Ясно, что он не имел никаких палеонтологических данных, подтверждавших его предположение, он базировался только на том факте, что в Африке в настоящее время живут два вида человекообразных обезьян - гориллы и шимпанзе. В "Происхождении видов" он пишет следующее:
"Следует доискиваться, где находится колыбель человеческого рода в том периоде его развития, в котором наши предки отделились от обезьяньего ствола. Факт, что человек относится к этому стволу, является доказательством того, что наши предки заселяли Старый Свет, а не Австралию или какой-нибудь океанический остров, как можно считать на основании закона, географического распределения.
В каждом обширном районе света живущие в настоящее время млекопитающие находятся в близком родстве с видами, развивающимися эволюционным путем. Поэтому весьма вероятно, что Африку раньше населяли вымершие обезьяны, бывшие в близком родстве с гориллой и шимпанзе, а так как эти оба вида в настоящее время являются наиболее близкими родственниками человека, можно принять, что наши давние предки жили в Африке, а не где-либо в другом месте. Однако было бы лишним дальше рассуждать на эту тему".
Позже начали принимать во внимание Азию в качестве предполагаемой колыбели человеческого рода. Это нашло свое основание в новых палеонтологических находках на территории Явы и Китая. Тем не менее, однако, как увидим ниже, Дарвин вероятно и в этом вопросе был прав.
Согласно Ф.Д. Дарлингтону (Ph.J. Darlington), американскому зоогео-графу, африканский континент был той территорией, на которой наступило отделение от животного ствола линии, ведущей к развитию человека. Эволюцию в направлении предчеловеческих форм до того момента, когда одно из племен человекообразных обезьян достигло вертикального положения тела, когда эти наши древнейшие предки начали ходить и бегать на двух ногах, освобождая этим передние конечности от функции передвижения. Сформировалась рука - орудие труда (Энгельс).
Некоторые из авторов считают, что изменение условий окружающей среды явилось непосредственной причиной изменения способа передвижения, который в свою очередь привел к далеко идущим преобразованиям всего организма, а прежде всего изменению в сочленении позвоночника с черепом, и преобразованию хватательного первого пальца стопы, в палец, лишенный этого свойства.
В результате охлаждения климата, густые тропические леса начали изреживаться, и обезьяны были вынуждены все чаще пребывать на земле, что привело к коренному изменению направления действия отбора. Отбор, используя наследственную изменчивость, привел в конце концов к развитию двуногих форм. Согласно этому взгляду, эволюционное развитие вертикального положения у наших предков было вызвано изменениями климата.
Ф. Дарлингтон (Darlington) придерживается другого мнения, и его взгляды, как нам кажется, заслуживают того чтобы к ним отнестись с пристальным вниманием. Согласно этому автору эволюционное формирование вертикальной позиции у наших давних предков произошло другим путем. Расширение ареала какой-то группы животных обычно не связано с бегством от невыгодных условий жизни, а наоборот, является проявлением активности этой группы, стремлением к использованию новых условий и заселению новых местностей.
"С самого начала - пишет Дарлингтон - обезьяны, которые со временем стали людьми, стремились к чему-то, а не убегали в опасении чего-то". По мнению этого автора, соответственные к тому условия существовали именно в Африке, где первобытные, мало специализированные человекообразные обезьяны, жившие в тропических лесах, устремились дальше на юг, в местности, лишенные уже густых тропических лесов.
Трудно предполагать, чтобы эволюция, идущая в направлении развития человека, могла начаться на территории северной Европы или Азии, так как там не было соответствующих условий. Кроме того формы эти должны были бы проникнуть также на американский континент через существовавшее тогда сухопутное соединение между Азией и Северной Америкой. Известно, однако, что в Новом Свете не найдено остатков человекообразных обезьян и первобытного человека. Поэтому наиболее вероятным является тот взгляд, что колыбелью человеческого рода была Африка.
Одновременно с принятием вертикального положения первобытный человек, живший в местностях, лишенных лесов, доставлявших его предкам обильную пищу, должен был и в других отношениях изменить образ жизни, который он вел до того времени. Современные обезьяны находят в густых лесах богатство разнообразной растительной пищи, а также яйца птиц, птенцов, мелких млекопитающих и многочисленных беспозвоночных. Этот образ питания должен был измениться у форм, живших на открытых местностях. Те, которые перед этим были сборщиками, превратились в охотников. Даже современные обезьяны бросаются случайно встретившимися ветками и камнями.
Эта способность приобрела особое значение, когда обезьяны поселились на открытых местностях. Умерщвление дичи на расстояние, а не пользование зубами в непосредственной борьбе, постепенно привело к атрофии клыков и резцов, то есть к постепенному изменению зубов на такие, какие имеет человек. Этот процесс эволюции животных в первобытных людей несомненно продолжался многие миллионы лет.
От времени превращения обезьян, живших на деревьях, в первые формы, способные к производству примитивных орудий, прошло десять, а может быть около двадцати миллионов лет. Таким образом, это был очень длительный период времени, который явился приготовительным к дальнейшей, уже протекающей в быстром темпе эволюции человека. Смотря на этот вопрос глазами геолога, следует принять, что эти первые, очень важные этапы эволюции, начались в миоцене, и продолжались дальше в течение плиоценовой эпохи.
Однако сохранились лишь скудные следы этого переломного периода нашей эволюции. Условия сохранения ископаемых остатков этой эволюционной линии в направлении человеческих форм были особенно неблагоприятными. Не сохранились также остатки первых орудий, так как это были предметы - ветви, камни, - не обработанные руками наших далеких предков. Поэтому невозможно отличить камней, которые могли служить в качестве примитивных орудий или оружия от других, которых не коснулась рука нашего предка. Несмотря на это палеонтология принесла ценные данные из третичного периода, которые остаются в согласии с взглядами, изложенными выше. Описываем эти находки на основании труда известного палеонтолога Л.С. В. Лики (L.S. W. Leakey).
Уже из олигоценовой эпохи известны в Египте остатки человекообразной обезьяны, Propliopithecus. Возможно, что эта форма или формы, сходные с ней, дали с одной стороны начало другим человекообразным обезьянам, известным из миоцена (Proconsul, Limnopithecus), как и другим формам Pongidae, которые обладали определенными признаками, приближающими их к современным человекообразным обезьянам. Propliopithecus можно, по крайней мере пока, считать исходной формой двух племен, которые в дальнейшем развились эволюционным путем в двух совершенно разных направлениях.
Большинство ископаемых форм Pongidae, открытых Лики (Leakey) и его сотрудниками в Африке (Кения) из миоценовой эпохи, относятся к роду Proconsul и Limnopithecus. У представителей рода Proconsul в котором систематики различают три отдельных вида, в строении нижней челюсти отсутствуют те специфические детали, которыми отличаются челюсти современных человекообразных обезьян. Таким образом, эта обезьяна не была так высоко специализирована, как современные Pongidae. Надбровные дуги на черепе одного из видов Proconsul (Proconsul africanus) не развиты так сильно, как у современных человекообразных обезьян.
Форма лобной кости напоминает строение лобной кости человека. Как вытекает из анализа фауны, современной виду Proconsul, эти животные жили на открытых просторах; передние и задние конечности их были почти одинаковой длины.
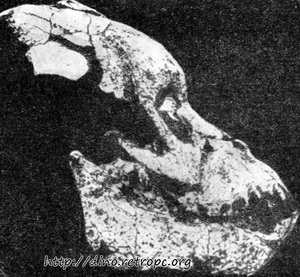
Рис. 71. Череп Proconsul africanus. Обращает на себя внимание отсутствие надбровных дуг; по П. Грассэ

Рис. 72. Реконструкция головы Proconsul; по Дж.Г. Симпсону и др.
Бесспорным фактом является то, что среди окаменелых представителей млекопитающих меньше всего находок относится к приматам. Эти животные жили или в лесах, или на открытых местах вблизи водоемов. Поэтому условия их жизни не способствовали образованию окаменелостей, то есть фоссилизации их костей. Процесс фоссилизации происходит тогда, когда остатки животного вскоре после его смерти будут смыты в воду и подвергнутся минерализации под воздействием воды, содержащей минеральные соли.
Несмотря на это, на основании приведенных до настоящего времени раскопок известно, что в миоцене и плиоцене Hominoidea были группой, богатой родами и видами, которые жили в Африке, Европе и Азии. В Европе и Азии жила форма Dryopithecus, а в Азии (Индии) жили еще и другие виды (Bramapithecus, Ramapithecus, Sivapithecus).
В Африке, однако, до настоящего времени не обнаружено никаких ископаемых форм представителей Hominoidea из плиоценовой эпохи. Наиболее ранние формы известны из нижнего плейстоцена. Прежде, чем мы перейдем к их описанию, следует в кратких чертах познакомиться с той частью кайнозойской эры, которую историческая геология называет четвертичным периодом.
Четвертичный период делится на две эпохи, а именно, на плейстоцен и голоцен, или современную эпоху. Продолжительность плейстоцена еще окончательно не установлена. Учитывая мнение многих авторов, следует принять, что он длился примерно пятьсот тысяч лет, а максимально около одного миллиона лет.
В плейстоценовую эпоху в северном полушарии, а также, хотя и в меньшей степени, в южном, наступило значительное похолодание и наблюдалось движение с севера массы льдов, достигших различной географической широты. В северном полушарии различаем четыре основных ледниковых периода, или периоды обледенения. Между каждыми двумя ледниковыми периодами наступали более теплые, межледниковые периоды, во время которых ледники таяли и отодвигались на север. Таким образом, различаем в плейстоцене на территории Евроазии четыре ледниковых периода и три более теплых, межледниковых периода. В Северной Америке авторы насчитывают пять ледниковых периодов.
Первый ледниковый период, так называемый гюнц, уступил место первому межледниковому периоду. После второго ледникового периода, минделя, наступил второй межледниковый период, а после третьего - рисса наступил третий межледниковый период. Наконец после третьего межледникового периода наступил четвертый ледниковый период - вюрм. После его окончания наступил современный период, то есть голоцен или аллювиум, продолжающийся уже около 20-25 тысяч лет.
Во время первого ледникового периода ледники охватили большую часть Польши, распространяясь в направлении России. Во время второго ледникового периода ледники распространились еще южнее. Третье обледенение было уже меньшим, а четвертое - наименьшим. Читателя, интересующегося этими вопросами, отсылаем к послевоенному изданию книги профессора А. Лота (Loth), "Человек прошлого", дополненной Мыдлярским.
В межледниковые периоды климат был более теплым, так что весь плейстоцен, называемый раньше дилювиальной эпохой, характеризовался очень большими климатическими колебаниями, что не могло не повлиять на процесс эволюции. Во время плейстоцена массы ледников занимали только часть северного полушария, то есть часть Европы Азии и Африки. Материки южного полушария подверглись гораздо меньшему охлаждению. Южная Европа и Азия, Африка и Южная Америка были свободны от внезапных изменений климата, и поэтому трудно произвести точное хронологическое сравнение окаменелостей северных и южных районов. С той же точки зрения, вместо того, чтобы выделять четыре ледниковых периода и три межледниковых, можно говорить о нижнем, наиболее раннем плейстоцене, среднем и верхнем, наиболее позднем плейстоцене.
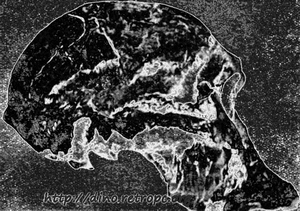
Рис. 73. Череп Австралопитека (Plesianthropus transvaalensis); по П. Грассэ
Период плейстоцена имеет особое значение для изучения эволюции человека, так как лишь в этом периоде мы находим остатки, относящиеся к формам, предшествующим появлению человека разумного (Homo sapiens). Из нижнего плейстоцена происходят очень интересные ископаемые формы, которые систаматика относит уже к семейству Hominidae.
Согласно некоторым авторам, семейство Hominidae, то есть людей, следовало бы разделить еще на два подсемейства: Australopithecinae и Нотiпiпае.
В 1925г. профессор Дарт (Dart) из университета в Иоганнесбурге описывает форму, найденную в южной Африке, которой дано родовое название Australopithecus. В дословном переводе это значит южная обезьяна, так как ископаемые остатки ее найдены на территории южной Африки. Это название послужило источником многих недоразумений. В настоящее время, имея кости, принадлежавшие многим особям разного возраста и пола, мы знаем, что австралопитеки не были обезьянами, а относились к семейству людей, хотя еще очень первобытных.
Вначале предполагалось, что найденные остатки относились также к концу третичного периода, плиоцену, однако позже, на основании анализа фауны, сопутствующей австралопитекам, было доказано, что наиболее ранние формы датируются началом плейстоцена.
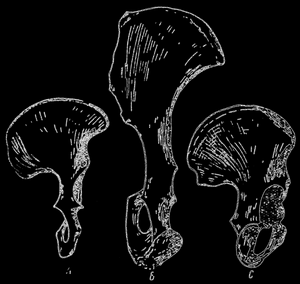
Рис. 74. Сравнение формы костей таза. А - австралопитек, В - шимпанзе, С - современный человек. Бедренная кость австралопитека и современного человека обладает некоторым сходством, бедренная же кость шимпанзе отличается продолговатой формой; по П. Грассэ
Палеонтологи обнаружили, что австралопитеки отличались уже целым рядом признаков, приближающих их к человеческому роду. Во первых, базируясь на строении таза, а прежде всего подвздошной кости, можно было установить, что эти формы передвигались на двух конечностях, то есть находились в вертикальном положении, и в результате этого верхние конечности, освободившиеся от функции локомоции, могли быть использованы для других целей.
Кроме того, зубная система австралопитека аналогична зубной системе, которую мы встречаем у человека. Зубная система является одним из тех признаков, которые изменялись незначительно и которые не зависят от качества пищи. (Это замечание автора не точно. Изменчивость зубной системы весьма значительна и зависит от пищи. Однако, несмотря на приспособительную изменчивость, особенности расположения основных бугорков зуба, их развитие и преобразование позволяют судить о степени родства ископаемых форм (прим. редактора).) Поэтому зубная система может являться хорошим показателем при оценке позиции исследуемых ископаемых форм в систематике.
Эти формы имели еще относительно небольшой мозг, близкий по величине к мозгу человекообразных обезьян, о чем можно судить по объему мозгового черепа. Согласно расчетам палеонтологов, объем мозгового черепа австралопитека колебался в пределах 450-700 см3, что более или менее соответствует емкости черепа гориллы (650 см3). Горилла является более крупным животным, чем был австралопитек. Во всяком случае эти формы, отнесенные в настоящее время к семейству Hominidae имели мозг лишь немногим больший, чем современные человекообразные обезьяны. Но сама величина мозга не предрешает его функциональной способности и можно принять, что австралопитек значительно превышал с этой точки зрения живущих в настоящее время Pongidae.
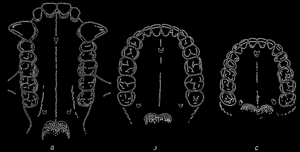
Рис. 75. Зубы верхней челюсти гориллы, австралопитека и современного человека (слева направо). Зубы верхней челюсти гориллы отличаются отдельным направлением специализации (линия зубов в виде буквы U, сильное развитие клыков, их расположение за линией зубов); по Ле-Грос-Кларку. (Le Gros Clark)
Некоторые ученые выделяют среди австралопитеков два и даже три отдельных рода, однако в настоящее время трудно сказать, является ли это вполне оправданным. Вообще в палеонтологической систематике господствует довольно большой хаос. Каждый из авторов, обнаруживающий новые ископаемые остатки форм первобытного человека, дает им отдельные родовые и видовые названия, хотя часто это не является обязательным и приводит к ненужному, искусственному расчленению обнаруженных форм.
Несомненно австралопитеки столь значительно отличались от находок, относящихся к хронологически более поздним формациям плейстоцена; что заслуживают особых родовых названий, а может следует даже отнести их к особому подсемейству.
Что бы мы однако ни сказали по этому поводу, уже сегодня не подлежит сомнению, что мы должны отнести их к семейству Hominidae. Австралопитеки обладали уже исключительно человеческой чертой, а именно, способностью к созданию культуры, а тем самым и к передаче ее.
Уже раньше Дарт, который открыл австралопитеков, сообщил об открытии примитивных каменных орудий, которые вероятно были сделаны их рукой. К тому же выводу склоняют нас и другие открытия. В расщелинах скал и гротах, в которых были найдены остатки австралопитеков, обнаружены также многочисленные черепа живших одновременно с ними павианов. Установлено, что из 52 животных 48 вероятно погибло в результате повреждений в области левой височной кости. Дарт предполагает, что австралопитеки наносили удары своим жертвам при помощи длинных трубчатых костей каких-то крупных животных.
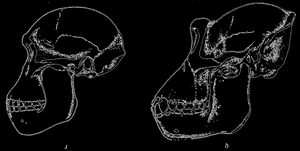
Рис. 76. Череп горилла (справа) и австралопитека (слева); по Ле-Грос-Клаоку
До недавнего времени, однако, мнение о том, что австралопитеки вырабатывали примитивные орудия, было лишь предположением. В настоящее время мы имеем уже убедительные доказательства этого.
В 1959 г. Лики (Leakey) вместе с женой открыли на территории Танганики череп, зубы и кость голени особи, которая относилась к австралопитекам и имела много признаков, сближающих ее с человеком. Эту форму Лики назвал Zinjanthropus boisei. Zinj по-арабски обозначает восточную Африку.
Лики следующим образом пишет о своем открытии: "Одним из наиболее бросающихся в глаза фактов в связи с Zinjanthropus является то, что череп их отличается признаками, свойственным роду Homo. Эти признаки до настоящего времени считались опознавательными признаками человека и некоторых вымерших человеческих форм, таких, как Pithecanthropus sinensis".
Так, например, у самцов гориллы и у некоторых австралопитеков сосцевидный отросток височной кости своей формой и величиной значительно
отличается от сосцевидного отростка человека. Тогда как у Zinjanthropus этот отросток не отличается от человеческого. Кроме того обнаружено, что представители этой формы австралопитека умели производить примитивные каменные орудия.
Zinjanthropus происходит из верхних слоев нижнего плейстоцена. Лики рассчитал, что должно было пройти 400000 лет или около 80000 поколении между Zinjanthropus и появлением человека. Автор считает, что Zinjanthropus был именно той формой среди австралопитеков, которая дала начало линии развития, ведущей к человеку и считает, что примерно полумиллиона лет достаточно на эволюционное преобразование Zinjanthropus в примитивного человека.
"В связи с этим - пишет Лики стоит напомнить, что если человек приручает какой-нибудь дикий вид животного, создается ситуация, в которой естественные процессы эволюции ускоряются, хотя темп мутации не изменяется. Когда человек начал производить орудия и начал контролировать свое окружение он тем самым стал "прирученным животным". Возможно, что преобразование в человека ускорило темп его морфологических изменений на ряду с быстрым развитием культуры".
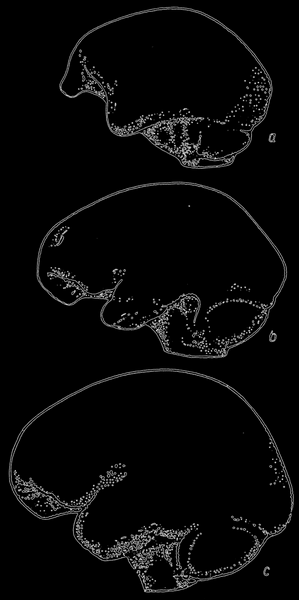
Рис. 77. Слепки внутренней поверхности черепа гориллы, питекантропа и современного человека (сверху вниз); по Ле-Грос-Кларку
В настоящее время мы располагаем интересными данными относительно темпа морфологических изменений, которые произошли в течение последних трехсот лет у зеленой обезьяны (Сеrсоpithecus aethiops sabaeus). Эти обезьяны живут в западной Африке. С давних времен негры привозили их на острова, относящиеся к Малым Антильским, где эти негры использовались в качестве рабов на плантациях сахарного тростника. На одном из островов (St. Kitts) часть этих обезьян живет в диком виде примерно с половины XVII века. У этих обезьян в течение 75-100 поколений наступили довольно большие морфологические изменения, касающиеся размеров черепа и зубов. Наблюдения показали, что темп изменений у этих представителей отряда приматов был гораздо более быстрым, чем изменений у представителей других отрядов млекопитающих, например, у лошадиных.
Замеченные изменения являются, по имению компетентных исследователей, генетическими изменениями, вызванными действием естественного отбора. - Эти исследования, как пишет Е.N. Ashton - указывают, что у высших приматов значительные изменения происходят во времени, которое с геологической точки зрения является бесконечно малым".
Теперь следует задуматься о том, в каком отношении находятся австралопитеки к нашему генеалогическому дереву? Большинство авторов считает, что род людской взял свое начало от форм, живших на территории Африки, которые относились к австралопитекам. Возможно, что именно такой прародительской формой является Zinjanthropus.
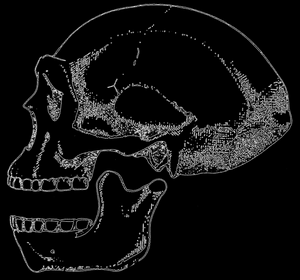
Рис. 78. Частично реконструированный череп Pithecanthropus pekinensis; по Ле-Грос-КларкуPithecanthropus pekinensis; по Ле-Грос-Кларку">
Согласно взглядом многих ученых, следующим этапом эволюционного развития человеческого рода являются формы, которые жили в среднем плейстоцене. Эти формы обнаружены на Яве и в Китае. Хронологически первые из этих находок имеют большое историческое значение.
В 1891 г. молодой голландский врач Дюбуа (Dubois), побужденный дискуссией о предполагаемой эволюции человека, которая тогда имела место, отправился на Яву и нашел там свод черепа и бедренную кость формы, которая по мнению нашедшего ее ученого и многих, современных ему дарвинистов должна была быть промежуточным эвеном между человекообразными обезьянами и человеком, то есть тем недостающим эвеном "missing link" в родословной Homo. Емкость черепа этой формы равнялась примерно 900 см3, то есть была средней между емкостью черепа современной человекообразной обезьяны и емкостью мозгового черепа современного человека. Поэтому Дюбуа назвал открытую им форму Pithecanthropus erectus, то есть обезьяночеловеком прямоходящим, так как строение бедренной кости несомненно свидетельствовало о вертикальном положении этой формы.
В течение длительного времени ничего не было известно о том, что Дюбуа позже обнаружил на Яве дальнейшие остатки тех же форм, которые, после смерти Дюбуа после второй мировой войны, стали доступны другим палеонтологам для исследований. Еще перед началом другой мировой войны голландец Кёнигсвальд (Koenigswald) нашел на Яве следующие окаменелости тех же форм.
Кроме того в 1924-1939 годах в пещерах Чжоу Коу Тьен вблизи Пекина было найдено 14 черепов особей разного пола и возраста, большое количество зубов, одиннадцать костей конечностей и разные лицевые кости. Хотя формы, найденные в Китае, были очень сходны с остатками, найденными на Яве, они получили особое видовое и родовое название. Китайские формы были названы Sinanthropus pekinensis.
Не подлежит сомнению, что формы из Китая и Явы относятся к одному виду, хотя их следует отнести к отдельным расам. Китайские формы имеют несколько большую емкость мозгового черепа. Кроме того установлено, что китайский Sinanthropus уже умел пользоваться огнем, о чем свидетельствуют находки в пещерах, где он жил. Он умел также делать каменные орудия. Возможно, что и формы, жившие на Яве, также умели пользоваться огнем и производили каменные орудия, которых, однако, до настоящего времени не удалось обнаружить. Все это указывает на то, что формы из Китая и Явы не были никакими человекообезьянами, а примитивными формами из рода Homo.
Однако различие между ними и более поздними формами человека слишком велики, чтобы их можно было отнести к тому же виду, что и человека. На этом основании некоторые из авторов, как, например, Добржанский, считают, что находки из Китая и Явы следует отнести к роду Homo, но к отдельному в этом роду виду, а именно, erectus. Таким образом, человек из Китая и Явы назывался бы Homo erectuse. Стремясь одновременно подчеркнуть различие между расой из Китая и Явы, первую называем Homo erectus pekinensis, а яванскую - Homo erectus erectus где третье название обозначает расу.
Палеонтологи считают, что в теплом климате Явы шансы на сохранение следов культуры этого первобытного вида человека были невелики. Это могло иметь место в холодном климате Китая, который заставлял этот древний вид людей искать убежища в пещерах. Все черепа, найденные в пещерах на территории Китая, были разбиты со стороны их основания, вероятно для того, чтобы вынуть мозг, а длинные кости конечностей были расколоты, вероятно для того, чтобы достать из них костный мозг.
Ввиду того, что в гротах не обнаружено ни позвонков, ни костей таза, можно предполагать, что Homo erectus был каннибалом. Своих жертв он четвертовал, а отрезанные головы и конечности переносил в свои пещеры.
Драгоценные китайские находки погибли во время второй мировой войны. За несколько недель перед началом войны между Америкой и Японией было согласовано, что ископаемые остатки, хранившиеся в американском учреждении, (Peking Union Medical College) будут перенесены в Америку. В начале декабря 1941 г. американский корабль на борту которого находились окаменелости Homo erectus, был затоплен вблизи Шанхая на реке Янцзыцзян. Остались, однако, точные слепки костных находок, выполненные палеонтологом Вейденрейхом (Weindenreich). В настоящее время в Китайской Народной Республике производятся дальнейшие исследования пещер и следует предполагать, что будут сделаны новые важные открытия.
Homo erectus многими морфологическими признаками отличается от вида Homo sapiens к которому относится современный человек. Homo erectus был невысокого роста, а его мозг, хоть и значительно больший, чем мозг австралопитека, был меньшим, чем мозг Homo sapiens.
Емкость мозгового черепа яванского человека равнялась 750-900 см3, а китайского 900-1200 см3 в сравнении со средней емкостью 1350 см3 современного человека. Кости черепа были более массивными, лоб покатый с выдающимися надбровными дугами, массивная нижняя челюсть с крупными зубами не имела характерного подбородка. Лицевой череп был сильно развит, что вместе с покатым лбом придавало ему животное выражение. Как уже было указано, зубы были крупными, хотя своей формой и строением соответствовали зубам человека. В некоторых черепах клыки несколько выступали над уровнем других зубов, а в верхней челюсти был небольшой промежуток между клыками и соседними с ними резцами.
Для эволюциониста наиболее важным является вопрос, имеется ли а если так, то какая, генетическая связь между австралопитеком и представителями вида Homo erectus. Морфологические исследования китайских и яванских раскопок свидетельствуют о том, что Homo erectus по своему строению стоял несколько ближе к человеку разумному, чем австралопитеки. Кроме того, он стоял на более высокой ступени культурного развития.
Австралопитеки жили в период, который хронологически соответствовал первому ледниковому периоду, а также, возможно, первому межледниковому. Homo erectus жил позже, так как остатки его относятся к первому межледниковому, второму ледниковому и второму межледниковому периодам. Во всяком случае так считают некоторые из палеонтологов. Все согласны с тем, что австралопитеки жили в периоде, предшествовавшем появлению Homo erectus что не исключает, конечно, возможности, что в течение некоторого времени эти формы могли существовать параллельно.
Одни из авторов считают, что Homo erectus был эволюционным звеном, объединяющим австралопитека с видом Homo sapiens, другие же предполагают, что хотя Homo erectus развился эволюционным путем из австралопитека, то он слишком дифференцировался в определенном направлении и вымер беспотомственно, а вид Homo sapiens развился из какого-то другого ствола австралопитеков. Следует подчеркнуть, что по мнению некоторых исследователей Homo erectus жил не только на территории Азии, но в северной Африке.
Так, например, Арамбур (Arambourg) относит форму, называемую Atlanthropus из северной Африки к формам, находившимся в близком родстве с Homo erectus. Если бы исследования подтвердили выводы Лрамбура, то есть если бы ареал распространения Homo erectus растягивался на Азию и Африку, многое говорило бы о том, что в нашем генеалогическом дереве следует считать Homo erectus звеном, связывающим австралопитека с формами, родственными виду, к которому относится современный человек.
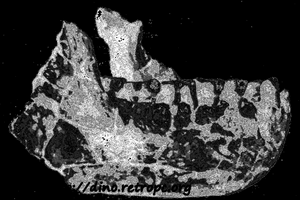
Рис. 79. Нижняя челюсть Atlanthropus mauritanensis, формы, находящейся в близком родстве с азиатским питекантропом, найденная в северной Африке; по П. Грассэ; по П. Грассэ">
Мы еще раз подчеркиваем, что разные названия, которыми названы разные формы, по крайней мере не свидетельствуют об их различии. Такие морфологические признаки, как строение скелета, обнаруживают большую изменчивость в популяциях людей. Несомненно и предки человека отличались большой изменчивостью. Отсутствие достаточного количества материала ископаемых форм не позволяет нам определить объема этой изменчивости.
Не следует забывать также о расовых различиях между разными группами первобытных людей, живших в разных местностях. Не только развитие человека разумного привело к образованию разных расовых различий, но, как мы указывали выше, Homo erectus встречался по крайней мере в двух разных расах.
Нам кажется, что наиболее трудно упорядочить целый ряд ископаемых форм, датирующихся более поздними геологическими периодами. Ископаемые остатки человека найдены во втором межледниковом периоде в Сванскомбе (Swanscombe) в Англии, в третьем ледниковом периоде в Steinheim в третьем межледниковом - в Эрингсдорфе (Ehringsdorf) и Fontechevade. Кроме того, как в третьем ледниковом, так и в четвертом ледниковом периоде живет форма, называемая неандертальским человеком.
В конце плейстоцена, то есть в период четвертого обледенения, появляется человек разумный, который как единственная форма живет до настоящего времени. Человек разумный, появившийся в четвертом ледниковом периоде, то есть примерно 75000 лет тому назад, обладает уже признаками, свойственными современному человеку, которого можно было бы назвать Homo sapiens sapiens, обозначая этим тройным названием его принадлежность к формам, живущим в настоящее время.
Наиболее трудным и спорным вопросом в палеонтологии является взаимоотношение Homo sapiens и Homo neanderthalensis. Мнения ученых по этому вопросу расходятся и нам кажется, что не скоро еще наступит согласование разных взглядов.
Для биолога приемлемой является концепция известного английского палеонтолога Ле-Грос-Кларка, а также замечания генетика Добржанского об эволюции человека. Вообще, как нам кажется, палеоантропология до настоящего времени не учитывает в достаточной степени успехов генетики, которые могли бы оказать большую помощь при установлении генезиса нашего рода.
Следует помнить, что современный человек, несмотря на деление на различные расы и популяции, представляет собой только один вид. Ни одна человеческая раса не является в отношении других генетически закрытой системой. Представители разных рас не только могут скрещиваться между собой, но в результате такого межрасового скрещивания потомство всегда бывает полностью плодовитым.
Хотя представители отдельных рас отличаются друг от друга многими соматическими признаками, объем изменчивости в каждой расе довольно велик. Поэтому, если антрополог имеет перед собой только части скелета, определение расовой принадлежности благодаря этой большой изменчивости часто является невозможным. Поэтому нет никакой уверенности в том, что уже в конце плейстоцена существовала дифференциация вида Homo sapiens на известные в настоящее время расы. Возможно, что лишь после конца четвертого ледникового периода в быстром темпе возникли известные в настоящее время расы современного человека.
Если, однако в настоящее время каждая из человеческих рас обладает такой большой изменчивостью, то наверняка и раньше еще в плейстоценовом периоде могли существовать разные расы. Ввиду меньшего количества населения и меньших возможностей скрещивания представителей разных популяций, шансы возникновения расовых различий были большими, чем в следующих периодах. Не следует также забывать, что ледниковые периоды также являлись фактором, облегчающим быстрое развитие генетического различия между человеческими популяциями, жившими в местностях, занятых ледниками, или вблизи них, и популяциями, жившими дальше на юг.
Можно согласиться со мнением Le Gros Clark что из вида Homo erectus постепенно, эволюционным путем, начали развиваться формы, которые отличались все большим сходством с Homo sapiens. К ним следует отнести формы, обнаруженные в Сванскомбе (Swanscombe), Штайнхайме (Steinheim), Фонтеш-ваде (Fontechevade) и другие.
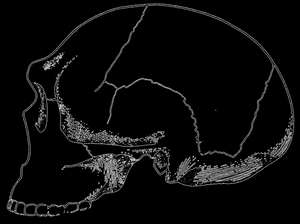
Рис. 80. Частично реконструированный череп из Steinheim. Хотя этот череп относится к неандертальским формам, он отличается среди них примитивным строением; по Ле-Грос Кларку
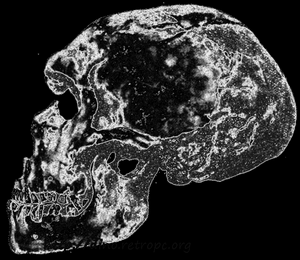
Рис. 81. Типичный неандертальский череп; по П. Грассэ
Общая эволюция человеческого рода происходила в направлении человека разумного. Однако эти формы, жившие в разных климатических условиях и значительной географической изоляции, отличались друг от друга. Главным образом в четвертом ледниковом периоде особые климатические условия, господствовавшие в Европе и Азии, способствовали обособлению формы, которая названа неандертальским человеком (Homo neandenhalensis). Типичные представители этих человеческих форм жили только во время четвертого ледникового периода на территории Европы, где и были раньше всего обнаружены. Родственные им формы, однако, встречаются уже в третьем межледниковом периоде.
По мнению многих исследователей, морфологические особенности скелета этих типичных неандертальцев так отличны от Homo sapiens, что их относят к особому виду. Отсюда и происходит название Homo neandenhalensis в противоположность Homo sapiens.
Приведем вкратце эти характерные особенности неандертальского человека, базируясь на определении этой формы, которое дает Ле-Грос-Кларк.
Череп прежде всего характеризуется массивными надбровными дугами. Лоб покатый. Лицевой череп хорошо развит. Массивная нижняя челюсть лишена характерного для человека разумного подбородка. Зубы более крупны, чем у Homo sapiens. Емкость мозгового черепа была большой и колебалась от 1300 до 1600 см3. Неандертальский человек был невысокого роста, коренастый, отличался большой физической силой, о чем свидетельствуют массивные кости и поверхности прикрепления мышц.
Классические неандертальские формы жили в Европе в четвертом ледниковом периоде, однако близкие им формы жили в Азии и Африке. Ввиду того, что человеческие и предчеловеческие формы отличаются большой изменчивостью, некоторые из современных авторов, а особенно генетики, считают, что несмотря на различия, которые имеют место в строении скелета, неандерталец был только расой, а не отдельным видом человека. В связи с этим большой интерес представляют находки ископаемых форм на горе Кармель в Палестине.
Вероятно эти находки относятся к формам, жившим в третьем межледниковом и в начале четвертого ледникового периода. Некоторыми своими признаками они приближаются к Homo sapiens, тогда как другими они сходны с неандертальцами. Некоторые из авторов предполагают, что палестинские формы являются продуктом скрещивания Homo sapiens с Homo neandenhalensis, другие зато считают, что эти формы представляют постепенный переход от форм человека неандертальского к человеку разумному.
Неандерталец жил в Европе еще 35-40 тысяч лет тому назад. Он жил в пещерах и на равнинах южной и восточной Франции группами в 30-50 человек. В холодном климате того времени в Европе жили разнообразные животные, как северные олени, песцы, зайцы, зубры, лошади, олени, мамонты, носороги, а из хищников - медведи, львы, гиены и волки.
Неандертальцы уже отличались высокой культурой, производили разнообразные каменные орудия, хоронили своих умерших, и, как можно считать, имели уже определенные религиозные верования. Пользовались естественными красителями для раскрашивания своего тела, но не оставили после себя никаких скульптур, ни картин, которые украшают пещеры Homo sapiens sapiens конца плейстоцена. Во всяком случае, как подчеркивает Bordes, профессор предъистории из Бордо, жизнь неандертальских племен не была такой, как иногда представляют неандертальцев - полулюдей, полуживотных, прячущихся в пещерах, запуганных и отличающихся низким уровнем интеллекта.
В конце плейстоцена неандертальцы погибли. Изменения климата привели к тому, что естественный отбор начал покровительствовать формам собственно Homo sapiens. По мнению многих исследователей, скрещивание неандертальских форм с формами собственно Homo sapiens закрепило во многих популя-циаях некоторые гены, относившиеся вначале к неандертальской расе. Это является еще одним доказательством, свидетельствующим о том, что неандертальские формы относились к генетически открытой системе, то есть не являлись отдельным видом.
Более подробный разбор раскопок, которые заслуживают внимания, так как на их основании можно было бы представить родословную человеческого рода, является скорей предметом антропологии. Однако уже на основании этих скупых данных, которые мы привели, можно, как нам кажется, сделать определенные общие выводы.
На рисунке 82 представлена родословная человека согласно Ле-Грос-Кларку. Это только очень схематическая попытка изобразить в общих чертах постепенное эволюционное развитие человеческих форм, которые произошли от животного ствола. Ясно, что новые открытия могут в значительной степени изменить приведенную схему. В настоящее же время новые находки не противоречат общим чертам приведенного эволюционного развития.
Человеческий род происходит из какого-то мало дифференцированного племени человекообразных обезьян. Он в основном прошел три главные ступени развития. Первую ступень представляли те формы, которые относились к австралопитекам. Они передвигались уже на двух конечностях, хотя мозг их был еще небольшим. Таким образом, вертикальное положение опережало в эволюционном процессе развитие мозга, которое отмечали на второй ступени развития, представленный Homo erectus, отличавшийся более высокой культурой, чем австралопитек.
Очень рано, примерно в половине плейстоцена, появляются формы, сходные в своих признаках с Homo sapiens. Эти формы образуют отдельные расы, из которых наиболее обособленной была неандертальская раса.
Как правильно подчеркивает Добржанский, эволюционное развитие человека отличается одной необыкновенно характерной чертой. Если в животном мире расовая дифференциация чаще всего приводила к возникновению многочисленных видов, живущих одновременно, то в филогенезе человеческих форм возникшие расы не достигали в одном и том же периоде времени уровня вида, и поэтому не существовало одновременно нескольких человеческих видов, родственных между собой. Возможно, что единственным исключением являлись два разные вида австралопитеков. Все данные свидетельствуют о том, что от момента преобразования австралопитека Homo erectus, он представлял один вид, хотя и разделенный на отдельные расы. Когда место Homo erectus занял Homo sapiens, все его расы само собой относились только к одному виду.
Ясно, что в течение некоторого, хотя и короткого времени, могли существовать параллельно два вида, старый и новый. Таким образом, в течение какого то времени кроме рода Homo мог еще жить какой-то вид австралопитека, предок Homo. Таким же образом, несмотря на появление Homo sapiens, могли еще в течение некоторого времени остаться группы, относящиеся к Homo erectus. Это видно и на схеме Ле-Грос--Кларка.
Можно, однако, предположить что старый вид, стоящий на более низком культурном уровне, быстро уступил свое место формам, относящимся к новому виду. Можно считать, что это свойство филогенеза человеческих форм не образовывать живущих параллельно многочисленных видов, развивающихся из общего ствола, было обусловлено новой особой способностью этих форм - создания культуры и общественного наследования. Благодаря этим особым свойствам человеческие формы не были изолированы друг от друга в течение достаточно длительного времени, что бы позволило на преобразование рас в отдельные виды.
Как обнаружил Лики, даже Zinjanthropus собирал камни, пригодные для изготовления примитивных орудий, с отдельных мест, а затем переносил на место своего жительства. Разные группы первобытных людей должны были быть в контакте между собой, и эти формы никогда не подвергались такой степени географической изоляции, как виды животных. Человеческие формы, именно благодаря своему психическому развитию быстро научились преодолевать изоляционные барьеры, стоящие на их пути.
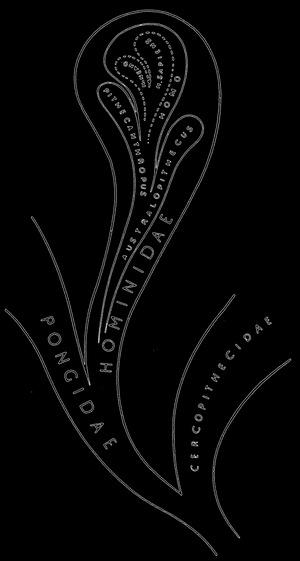
Рис. 82. Схематическое изображение филогенетических взаимоотношений среди Hominidae; по Ле-Грос-КларкуHominidae; по Ле-Грос-Кларку">
Как мы уже указывали, в приведенной родословной человека могут еще наступить многочисленные изменения. Нельзя забывать, что условия не способствовали сохранению до сегодняшнего дня более многочисленных окаменелостей, которые, кроме того, имеют лишь фрагментарный характер. Только ввиде исключения обнаружены остатки, относящиеся к большому числу особей, скелеты которых относительно хорошо сохранились. Мы имеем особенно мало данных, касающихся самого эволюционного развития человекообразных обезьян.
Очень важными для расшифровки нашей эволюции являлись бы находки с самого конца плиоцена. Однако общие черты нашей родословной так хорошо изучены, что "выдающееся" открытие в Пильтдауне (Piltdown) в Англии заставило сразу усомниться в их подлинности. Выглядело это следующим образом.
В 1912г. и в следующие годы в Пильтдауне были найдены череп и нижняя челюсть, которые будто бы принадлежали одной особи. Если череп, несомненно, принадлежал к Homo sapiens, то нижняя челюсть имела отчетливые признаки челюсти какой-то человекообразной обезьяны. Это странное объединение в одной особи человеческих и обезьяньих признаков склонило многих палеонтологов к выводу, что не стоит уделять ей внимания, так как с точки зрения палеонтологии такое создание не могло существовать.
Однако лишь в 1953 г. английские ученые, пользуясь новой методикой, сумели установить, что кости, найденные в Пильтдауне кто-то необыкновенно хитро подогнал искусственно. Другими словами, остатки формы, названной Eoanthropus dawsoni оказались фальсификацией.
Этот случай является настолько поучительным, что если вначале открытие в Пильтдауне вызвало большое волнение в мире ученых, то позже, после более тщательного анализа, им no-существу перестали интересоваться. Оно не укладывалось в открытых главных линиях развития человека. Оказалось, что взгляды палеонтологов были правильными. Eoanthropus не мог существовать и действительно его никогда на было.